Правдин И.Ф.
Руководство по изучению рыб (преимущественно пресноводных)
Четвертое издание переработанное и дополненное
Под ред. проф. П.А. Дрягина и канд. биол. наук В.В. Покровского
Издательство «Пищевая промышленность» Москва 1966 г.
Предисловие
Книга проф. Й. Ф. Правдина «Руководство по изучению рыб» с 1926 по 1939 г. выдержала три издания. Учитывая массовые запросы на эту книгу, И. Ф. Правдин готовил к печати четвертое, значительно переработанное и дополненное издание. Но довести до конца эту работу ему не удалось, 23 февраля 1963 г. Иван Федорович скоропостижно скончался.
Труд по окончанию подготовки к печати книги И.Ф. Правда взяли на себя его товарищи по совместной работе и ученики.
Проф. П. А. Дрягин (ГосНИОРХ) сделал ряд дополнений к «Введению», в частности в определение содержания ихтиологии, составил перечень важнейших задач ихтиологических исследований. Им же написаны разделы: «Изучение жизненных циклов рыб» к главе VI и «Методы определения скрытых (потенциальных) возможностей роста рыб» к главе VIII.
Кандидатом биологических наук Г. А. Амирхановым сделано дополнение к главе II-«Применение иммуносерологических реакций в ихтиологии».
Г. Г. Галкиным (ГосНИОРХ) дополнены главы V, VII и VIII. Член-корреспондент АН Латвийской ССР проф. Я. Я. Лус дополнил главу XII.
В. В. Покровским (ГосНИОРХ) расширен обзор основной литературы по методике ихтиологических исследований и о рыбах водоемов СССР в главе І. Им же составлен раздел – « Популяционная плодовитость» к главе IV, переработана глава IX написаны к главе Х разделы: «Примеры определения фактической численности рыб, ихтиомассы и возможных уловов в обработанных химическим методом озерах» и «Математическое моделирование динамики численности рыб». Раздел этой главы « Краткий обзор методов определения рыбных запасов и прогнозирования уловов» дополнен проф. П. В. Тюриным и В. В. Покровским кратким изложением методики П. В. Тюрина об определении коэффициента естественной смертности рыб (книга проф. Тюрина вышла в 1963 г. и не могла быть использована Й. Ф. Правдиным, так же как и работы Е. В. Бурмакина о химическом методе рыбохозяйственного преобразования озер и В. В. Меншуткина об опыте моделирования динамики численности популяций рыб).
Л. П. Тихомирова (ГосНИОРХ) уточнила и дополнила литературу, использованную при составлении «Руководства», которую редакция сочла целесообразным вынести из текста в отдельный список.
Следует отметить, что И. Ф. Правдиным выполнена очень большая работа по подготовке нового издания «Руководство по изучению рыб». Большое участие в окончательной подготовке рукописи к печати приняла супруга автора-М. А. Правдина.
Можно рассчитывать, что книга И. Ф. Правдина – выдающегося советского ученого и прекрасного педагога – принесет большую пользу ихтиологам и особенно начинающим исследователям, для которых в основном и предназначал Й. Ф. Правдин свою книгу.
Редакция
Введение
После выхода в свет нашего «Руководства по изучению рыб», изданного в 1939 г. Ленинградским государственным университетом, прошло 25 лет. За этот период проведено много ихтиологических экспедиций на морях, водохранилищах, озерах и реках. Возникли новые учебные и научно-исследовательские учреждения, занимающиеся вопросами рыбного хозяйства. Советская ихтиологическая наука решила много вопросов, важных для развития нашего рыбного хозяйства, но предстоит еще решить много проблем.
Содержание ихтиологии и положение ее среди сопредельных наук, в основном сформулированные Л. С. Бергом и изложенные в статье Е. Н. Павловского (1953), можно представить в следующем виде.
Ихтиология (от ichthyos-рыба и logos-учение) изучает внешние признаки и внутреннее строение рыб (гистологию, морфологию, анатомию), жизнедеятельность рыб (питание, размножение, миграции, возраст, рост), функционирование различных органов (физиологию и биохимию), отношение рыб к внешней среде-неорганической и органической и установление закономерностей динамики численности рыб (экологию), индивидуальную историю развития (онтогенез) и историю развития видов, ;родов, семейств, отрядов и т. д. (эволюцию или филогенез), систему рыб ныне живущих (систематику) и ископаемых (палеонтологию), наконец, географическое распространение и видовой состав рыб отдельных областей (зоогеографию).
Таким образом, ихтиология – наука, всесторонне изучающая рыб и имеющая множественные связи с другими науками.
Исходя из современных требований обеспечения населения Советского Союза изобилием высококачественных рыбных продуктов, можно определить следующие важнейшие задачи дальнейших ихтиологических исследований (по Дрягину, 1962):
а) разработка методов рыбохозяйственной оценки расового состава отдельных стад промысловых рыб на морфологической, биологической, экологической и биохимической основах и организация племенной работы на рыбоводных заводах, рыбопитомниках, нерестово-выростных хозяйствах;
б) разработка проблемы цикличности жизненных явлений у рыб в онтогенезе и филогенезе; исследование жизненных циклон рыб не только по отдельным фазам, этапам и стадиям индивидуального развития, но и в их взаимозависимости, а также изучение жизненных циклов в целом с учетом динамики численности стад в связи с различными условиями обитания; исследование половых циклов рыб и их преобразования в зависимости от разных условий обитания;
в) разработка основ направленного изменения состав ихтиофауны, формирования размерной, возрастной и половой структуры популяций рыб в рыбохозяйственных водоемах; определение оптимальной плотности стад рыб, оптимальных промысловых размеров (руководствуясь при этом тщательной оценкой добываемой рыбы как пищевого продукта) и рационального регулирования рыболовства, обеспечивающих повышение рыбопродуктивности водоемов;
г) установление закономерностей суточных и сезонных жизненных ритмов, а также суточного и сезонного распределения рыб в водоемах;
д) усовершенствования методики ихтиологических исследований с внедрением новых технических средств.
Односторонние исследования уступили место исследованиям комплексным, когда каждый существенный вопрос одновременно решается различными специалистами. Современный научный работник-ихтиолог должен владеть познаниями и методами не только «своей» узкой специальности, но и других дисциплин, которые могут содействовать всестороннему освещению исследуемого вопроса.
Необходимо сопоставлять одно явление с другим, находить причинность и закономерность явлений.
В Советском Союзе ихтиологическими исследованиями занимаются союзные и республиканские научно-исследовательские институты морского (ВНИРО), озерно-речного (ГосНИОРХ) и прудового (ВНИИПРХ, Украинский институт) рыбного хозяйства и их многочисленные отделения, Академия наук СССР, республиканские Академии наук, государственные университеты и другие высшие учебные заведения, географические и краеведческие общества и др. В исследованиях рыб участвует много ихтиологов, рыбоводов, гидробиологов и других специалистов. На эти исследования Советским правительством отпускаются колоссальные средства.
Многолетний опыт автора в чтении курса методики ихтиологических исследований в Ленинградском и Петрозаводском университетах, а также занятия в научно-исследовательских учреждениях и руководство аспирантами показывают, что ихтиологи в начале своей научной деятельности нуждаются в пояснениях и обоснованиях методических приемов по разным вопросам ихтиологии, которые и даны в настоящем Руководстве.
Несмотря на значительные дополнения, новое издание Руководства не свободно от недостатков. Трудно пока рекомендовать например, бесспорную методику определения запасов рыб, поскольку этот вопрос продолжает быть дискуссионным и решается разными авторами по-разному.
Основное содержание книги относится к методике исследований пресноводных рыб, но некоторые из методических рекомендаций приложимы и к исследованиям ихтиофауны морских водоемов (разделы по изучению видового состава и биологии рыб, по изучению возраста и роста, по применению вариационной статистики и др.).
Наша книга предназначается в основном для начинающих ихтиологов.
Выпуску в свет нового издания мы обязаны многим запросам со стороны научно-исследовательских и рыбохозяйственных организаций, учебных заведений и отдельных научных работников.
Автор особенно благодарен П. А. Дрягину и В. В. Покровскому, которые сказали большую помощь в составлении этой книги.
Автор Январь 1963 г.
Глава І
ОСНОВНАЯ ЛИТЕРАТУРА ПО МЕТОДИКЕ ИХТИОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ И РЬІБАМ ВОДОЕМОВ СССР
По ряду разделов ихтиологической методики и методов рыбохозяйственных исследований имеются хорошие и достаточно полные сводки. Рассмотрим следующие из них.
В книге Л. С. Берга «Рыбы пресных вод СССР и сопредельных стран» (1948, 1949) дано много методических указаний об определении морфологических признаков рыб, имеющих систематическое значение. Излагаются понятия о виде и внутривидовых подразделениях в ихтиологии, а также приведены сведения о географическом распространении рыб Голарктики.
В 1949 г. под редакцией академика Л. С. Берга, А. С. Богданова, Т. С. Расса, Н. И. Кожина выпущен «Атлас промысловых рыб СССР». К атласу приложен текст с описаниями рыб, который облегчает начинающим исследователям определение рыб.
В 1940 г. Л. С. Бергом опубликована монография «Система рыбообразных и рыб, ныне живущих и ископаемых», переизданная в 1955 г. В этой работе, имеющей мировое значение в ихтиологии, дается наиболее совершенная для настоящего времени характеристика всех высших таксономических единиц рыбообразных и рыб.
Ихтиологам полезно познакомиться с книгами Э. Майра «Систематика и происхождение видов» (1947) и Э. Майра, Э. Линсли и Р. Юзингера «Методы и принципы зоологической систематики» (1956). В этих сводках разбираются вопросы систематики животных, в том числе и рыб, особенно их внутривидовых подразделений.
Много нового в сложный вопрос о внутривидовой неоднородности рыб внес Н. В. Лебедев в работе об элементарных популяциях у рыб (1946), а также П. А. Дрягин (1949), обосновавший деление рыб на биоэкологические группы. Внутривидовые
группировки у осетровых выделяет Н. Л. Гербильский (1950, 1953). О проблеме симпатрического видообразования есть полезная статья Б. М. Медникова (1963).
Половые циклы, плодовитость, размножение и жизненные циклы рыб рассматриваются в работах П. А. Дрягина (1939. 1949, 1952 и др.), В. С. Ивлева (1953), Б. В. Кошелева (1961), С. Г. Крыжановского (1953), Г. В. Никольского (1953 и др.), Б. Г. Иоганзена (1955 и др.), Ю. Г. Юровицкого (1958), И. Ф. Правдиным (1958) составлен обзор методов определения плодовитости рыб.
Морфо-экологическому исследованию рыб на ранних стадиях развития посвящены работы В. В. Васнецова и С. Г. Крыжановского и их сотрудников и учеников, опубликованные главным образом в Трудах института морфологии животных АН СССР им. А. Н. Северцова.
Статьи иллюстрированы превосходными рисунками, характеризующими морфологические особенности молоди сельдевых, сиговых, щуковых, карповых, окуневидных и других рыб.
Результаты исследования развития рыб Амура изложены в работах Б. В. Веригина, С. Г. Крыжановского, А. И. Смирнова, С. Г. Соина и других ихтиологов (труды Амурской ихтиологической экспедиции, 1950-1952).
Существенное значение имеет выполненная под руководством В. В. Васнецова и изданная АН СССР в 1948 г. работа «Морфологические особенности, определяющие питание леща, воблы и сазана на всех стадиях развития».
Т. И. Привольневым (1959) составлено обстоятельное руководство по методам исследования развития рыб.
П. А. Дрягин отмечает, что за последний период выполнено много ценных исследований по изучению жизненных циклов рыб и что дальнейшая задача заключается прежде всего в изыскании способов воздействия на разные и, особенно на ранние стадии развития рыб, которые содействовали бы увеличению выхода ценных рыбных продуктов и ускорению оборота хозяйства (Дрягин, 1961).
Лучшей книгой для начинающих заниматься изучением возраста рыб является «Руководство по изучению возраста и роста рыб» Н. И. Чугуновой (1959). В этой книге подробно изложены все современные способы определения возраста и роста рыб.
Хорошим пособием для ихтиологов является «Атлас чешуи пресноводных костистых рыб» Г. Г. Галкина.
О возможном (потенциальном) росте рыб- вопросе весьма существенного теоретического и практического значения – писали Е. К. Суворов (1940), П. А. Дрягин (1947, 1948).
Разработкой методов изучения питания рыб занимались многие исследователи. Специальные руководства, инструкции и методические статьи по этим вопросам написаны В. Г. Богоровым (1934), Е. Н. Боковой (1955), Е. В, Боруцким (1955), В. А. Броцкой (1939), Т. Ф. Дементьевой и Б. С. Ильиным (1938.), В.С. Ивлевым (1955), В. И. Жадиным (1956, 1960), М. Н. Лишевым (1950), П. Л. Пирожниковым (1953), К. Р. Фортунатовой (І955), А. А. Шорыгиным (1946, 1948, 1952) И.А. Щербининым (1955) и др. В 1961 г. Академией наук СССР издано обстоятельное «Руководство по изучению питания в естественных условиях», составленное ведущими специалистами в этой области под общей редакцией Е. В. Боруцкого. В руководстве подробно разбирается методика изучения пищи планктоноядных, бентосоядных, растительноядных и хищных и рыб, дается список основной литературы.
Методическое значение имеет работа В. А. Амосова по изысканию новых показателей экстерьера и упитанности рыб. Им опубликована статья «О новом показателе упитанности рыб – индексе удельной вальковатости» (1961) и другие работы по этому вопросу.
В 1947 г. вышла в свет прекрасная монография П. Ю. Шмидта о миграциях морских, проходных и пресноводных рыб и методах их изучения. Специальные указания о способах мечения рыб даны в инструкции Г. А. Караваева (1938, 1958). Начато применение метода мечения рыб радиоактивными индикаторами. Этому вопросу посвящены статьи А. С. Трошина (1956) В. С. Кирпичникова, А. Н. Световидова и А. С. Трошина (1958) И. А. Шехановой (1958) и др.
Ю. Ю. Марти (1948) принадлежит весьма обстоятельная сводка о методах промысловой разведки рыбы. Много статей по атому вопросу опубликовано в периодических изданиях «Рыбное хозяйство», «Вопросы ихтиологии» и других, в частности (быстро развивающихся сейчас методах подводных исследований (Зайцев и Радаков, 1960; О. Н. Киселев и Б. С. Соловьев 1961 и др.). В 1958 г. ВНИРО был издан специальный сборник «Поведение рыб и промысловая разведка».
Проблеме определения численности рыб и прогнозированию уловов посвящена книга С. В. Аверинцева (1948), а также ряд капитальных работ Г. Н. Монастырского (1952, и др.) и статья П. А. Дрягина (1951). В 1952 г. состоялась Всесоюзная конференция по вопросам рыбного хозяйства, а в 1960 г.- совещание по динамике численности рыб. В Трудах конференции (1953) и совещания (1961) єесть много ценнейших обобщений и предложений по совершенствованию методов разработки 5той сложнейшей проблемы. Ей уделяется большое внимание и в книге П. Г. Борисова и А. С. Богданова «Сырьевая база (рыбной промышленности СССР» (1955). В 1961 г. Академией наук СССР изданы «Очерки по биологическим основам рыбного хозяйства», где помещены статьи по теории динамики численности промысловых рыб.
П. В. Тюрин в книге «Биологические обоснования регулирования рыболовства на внутренних водоемах» (1963) предлагает метод определения коэффициента естественной смертности рыб. В этой книге доны указания о методах сборов и обработки ихтиологических материалов.
В последние годы рыбохозяйственная наука получила возможность определять абсолютную численность рыб и ихтиомассу в небольших озерах благодаря разработанному и внедряемому Е. В. Бурмакиным методу химической обработки водоемов (Бурмакин и Жаков, 1961; Бурмакин, 1963). Г. В. Никольский (1963) разработал биологические основы принципа математического моделирования в ихтиологии, а В. В. Меншуткин (1964) выполнил превосходный опыт исследования динамики численности рыб на основе представления популяции в виде кибернетической системы.
Обобщающие сводки по акклиматизации ценных рыб и выращиванию их в различных водоемах составлены Б. Г. Иоганзеном и А. Н. Петкевичем (1946), П. А. Дрягиным (1953, 1956), Е. В. Бурмакиным (1963), А. Ф. Карпевич (1960). Этому вопросу было посвящено и специальное Всесоюзное совещание, итоги которого опубликованы в «Трудах совещания по проблеме акклиматизации рыб и кормовых беспозвоночных» (1954).
В 1962 г. Академией наук СССР издано «Руководство по методике исследований физиологии рыб» и книга Н. С. Строганова «Экологическая физиология рыб». Издательством иностранной литературы в 1953 г. выпущен сборник статей по биохимии рыб (перевод с английского).
Выдающимся трудом является монография Ю. Г. Алеева (1963) «Функциональные основы внешнего строения рыбы». В книге рассматриваются приспособления рыб к движению, маскировке, захватыванию пищи и пр. Установленные автором особенности и закономерности строения рыб, связанные с их движением, уже используются, например, при конструировании плавательных аппаратов.
Много методических указаний по разным разделам рыбохозяйственной науки содержат книги Г. В. Никольского «Биология рыб» (1944) и «Экология рыб» (1963), а также известное руководство Е. К. Суворова «Основы ихтиологии» (1948).
Много работ, ценных в методическом отношении, публикуется в журналах «Вопросы ихтиологии», «Рыбное хозяйство», «Рыбоводство и рыболовство» и в периодических изданиях, выпускаемых научно-исследовательскими рыбохозяйственными учреждениями. Основные из них: Труды Всесоюзного научно-исследовательского института морского рыбного хозяйства и океанографии (ВНИРО) и Известия государственного научно-исследовательского института озерного и речного рыбного хозяйства (ГосНИОРХ, ранее ВНИОРХ). Периодические издания публикуют также филиалы и отделения этих институтов. Вопросы методики ихтиологических исследований освещаются также в изданиях университетов, педагогических институтов, академических учреждений, географических и краеведческих организаций и др.
Периодические информации об опубликованных работах по ихтиологии и вопросам рыбного хозяйства помещаются в журнале «Рыбное хозяйство», а также (с краткими аннотациями) в «Реферативном журнале». Есть указатели литературы по отдельным бассейнам, составленные В. К. Есиповым (1935) и Н. С. Романовым (1955, 1959).
В иностранной ихтиологической литературе за те же 25 лет опубликовано немало полезных для методики материалов, некоторые зарубежные методические источники последних лет использованы в настоящей книге. Но первое место в разработке методики принадлежит советским ихтиологам.
Ихтиологические исследования по отдельным промысловым бассейнам рассматриваются в следующих обзорах: Л. С. Берг «Очерк промысловых исследований в России» (1924); обзоры ихтиологических исследований за период с 1917 по 1937 г. опубликованы Л. С. Бергом (1937), а также Г. У. Линдбергом (1937) и Г. В. Никольским (1938). В 1945 г. Л. С. Берг и А. Н. Световидов сделали обзор достижений советской ихтиологии за 1917-1942 гг. В 1949 г. вышла в Ученых записках Карело-Финского университета моя статья об ихтиологических работах за время 1917-1947 гг. П. Г. Борисовым (1964) написан исторический очерк научно-промысловых ихтиологических исследований морских и пресных водоемов СССР, П. Г. Борисовым и Г. В. Никольским (1961)-обзор отечественных рыбохозяйственных исследований за 100 лет.
Основная литература по промысловым водоемам указана Е. К. Суворовым в его книге «Промысловые водоемы СССР» (1948).
Лучший определитель для пресноводных рыб-уже упоминавшаяся книга Л. С. Берга «Рыбы пресных вод СССР и сопредельных стран». Там есть и описания многих морских рыб, например рыб Каспийского моря. Широким распространением пользуется «Определитель промысловых рыб СССР» П. Г. Борисова и Н. С. Овсянникова. П. Й. Жуков (1960) составил краткий определитель рыб водоемов Белоруссии, им же дано описание рыб бассейна Немана (1958). В 1951 г. вышла книга Н. П. Танасийчука «Промысловые рыбы Волго-Каспия», в 1963 г.-Е. Н. Казанчеева «Рыбы Каспийского моря», в 1964 г. А. Н. Световидова-«Рыбы Черного моря».
Г. В. Никольским (1940) составлена обстоятельная монография по рыбам Аральского моря. Им же даны сведения обо всех видах рыб СССР в учебнике «Частная ихтиология».
Рыбы дальневосточных морей, описаны В. К. Солдатовым и Г. У. Линдбергом (1930). Кроме того, имеется хороший краткий определитель рыб советского Дальнего Востока и прилежащих, вод А. Я. Таранца (1937), П. Ю. Шмидт составил обзоры рыб Тихого океана (1948) и Охотского моря (1950). Карельским книжным издательством выпущена книга «Рыбы Белого моря» (1958). Рыбы северных морей СССР описаны А.П. Андрияшевым (1954).
Опубликован ряд сводок о рыбах отдельных промысловых районов и озерно-речных бассейнов СССР.
Л. С. Бергом (1940) дан обзор рыб Финского залива, Л. С, Бергом и И. Ф. Правдиным (1948)-рыб Кольского полуострова, Карельским отделением ГосНИОРХ в 1959 г, выпущен справочник «0зера Карелии», в котором помещена обзорная статья В. В, Покровского и П. И. Новикова об озерах и пресноводных рыбах Карелии. И. Ф. Правдиным (1954) составлена монография о сигах Карелии, П. И. Новиковым (1953) написан очерк об атлантическом лососе (семге), В. Г. Мельянцевым о форелях (1952) и палиях (1953), В, В, Покровским (1953) о ряпушках Карелии, Е. В. Бурмакин (1953), а также Г. А. Головков и А. Н. Кузьмин (1963) описали пелядь, И. С. Титенков (1961)– кубенскую нельму, Л. А. Кудерский (1958, 1964) -судака.
И. И. Николаевым (1953) дан перечень рыб Латвийской ССР, А. И. Амброзом (1956) описаны рыбы бассейна Днепра, С. Н. Троицким (1948) рыбы Краснодарского края.
Имеется подробное описание рыб р. Лены (Борисов, 1928) и водоемов Якутии (Дрягин, 1933). 0 промысловых рыбах Обь-Иртышского бассейна обстоятельная сводка дана П. А, Дрягиным (1948), а также. Б. Г. Иоганзеном (1948) и Б. К. Москаленко (1958). Г. В, Никольским и др. (1947) описаны рыбы верхней Печоры.
В 1955 г, опубликована прекрасная монография Д. Н. Талиева о бычках-подкаменщиках Байкала, а в 1958 г, вышла большая книга «Рыбы и рыбное хозяйство в бассейне озера Байкал», составленная коллективом под редакцией М. М. Кожова и К. И. Мишарина.
Ценные ихтиологические работы изданы Московским обществом испытателей природы о рыбах Амура (1950, 1951). Издательством Академии наук СССР выпущена книга Г. В. Никольского «Рыбы бассейна Амура» (1956). 0 рыбах Киргизии написал В. А. Турдаков (1963), о рыбах Таджикистана–Г. В. Никольский (1938). Обзор рыб Грузии и Армении дал Г. П. Барач (1940, 1941), Ю. А. Абдурахманов (1962) описал рыб пресных вод Азербайджана.
Глава 11
ИССЛЕДОВАНИЯ СИСТЕМАТИКИ РЫБ
Таксономические единицы и правила научной номенклатуры
Академик Л. С. Берг создал совершенную систему рыб, в которой генетическое родство отдельных классов, отрядов и семейств представлено с возможной полнотой (Берг, 1940, 1948–1949, 1955). Она дает обширнейшую тематику, громаднейшие перспективы для работы других исследователей (Правдин, 1945). Более полной и естественной системы рыб пока не предложено.
При работах по систематике рыб следует руководствоваться теми определениями таксономических единиц, какие даны Л. С. Бергом в 1 части его книги «Рыбы пресных вод СССР» (1948). По той же книге ихтиолог должен изучить правила научной номенклатуры. Исполнение этих двух требований (о таксономических единицах и номенклатуре) поможет начинающим ихтиологам избежать многочисленных трудностей и ошибок.
Таксономические единицы Л. С. Берг делит на две категории:
А – единицы географические и Б – единицы негеографические.
К первым он относит вид, подвид и племя.
Видом (species) Берг называет «совокупность особей, занимающих определенную географическую область и обладающих рядом определенных признаков, передаваемых по наследству и всегда отличающих данный вид от близких видов».
Подвид (subspecies)–совокупность особей, связанных с близким к ней видом наличностью переходных форм, но имеющая определенную область обитания, Подвид обладает одинаковой с видом стойкостью в передаче наследственных свойств,
Племя (natio) Л. С. Берг понимает как подвид второго порядка. Племя объединяет группу особей, связанных с подвидом переходными формами, имеющую наследственные признаки и определенное место распространения (ареал),
Ко второй (Б) категории таксономических единиц Л. С. Берг относит: расу или элементарный вид, экотип или инфраспециес (infraspecies)и морфу (morpha).
Расу он характеризует как группу стойких форм, связанных с видом переходными признаками, но oтличимыx по комбинации признаков. «0блacть распространения расы географически не обособлена от ареала вида; иногда расы бывают приурочены к определенным экологическим условиям – глубине, температуре воды, времени нереста, грунту и пр. раса обычно передает свои признаки по наследству и при изменении внешних условий», На расы распадаются не только виды, но и подвиды. Расы подвида Л. С. Берг предлагает называть инфpacyбcпeциec (infrasubspspecies). Еще более мелкая таксономическая единица, чем раса – биoтип.
H. B. Лебедев (1946) вводит понятие об элементарных популяциях рыб, понимая под таким термином группу рыб, пространственно обособленную и состоящую из многочисленных особей, имеющих одинаковый жизненный цикл. Элементарная популяция, по мнению Лебедева, – биологическая группа.
Mopфa, по Л. C. Бергу, – «более или менее резкое уклонение основного вида или подвида, охватывающее обширные группы особей или периодически целое поколение и вызванное резким изменением некоторых специальных условий существования, например: субстрата, пища и т. п., а иногда, быть может, зависящее от внутренних причин», морфы «не имеют географически определенной области обитания, встречаясь спорадически совместно с основной формой». Признаки морфы неустойчивы: «при перемене внешних условий морфа часто превращается в первобытное состояние.
Г. B. Hикoльcкий (1954) в понятие о виде вводит такое содержание, «Вид характеризуется относительной морфологической стабильностью, являющейся результатом приспособления к определенной среде, в которой вид формировался и живет. Изменчивость в пределах вида не выходит за границы его морфологической специфики, признаки вида отражают приспособление к определенным условиям существования, Вид занимает определенную область распространения, в пределах которой условия существования соответствуют его морфологическим особенностям.
Вид относительно стабилен во времени, т.е. раз возникнув, он сохраняет свою морфологическую специфику в течение всей своей истории… Историческое развитие идет не путем постепенного изменения существующих видов, а путем образования новых видов в результате изменения взаимоотношений особей, слагающих вид, и окружающих условий существования, т. е. видообразование, есть результат изменения условий существования и связанных с ним морфологических особенностей популяции, путем перестройки организмов и среды».
Из правил научной номенклатуры укажем наиболее существенные. Виды обозначаются бинoминaльнo, т. е.· название вида должно включать наименования: родовое (название рода пишется всегда с заглавной буквы и обычно имеет греческое словопроизводство) и видовое (пишется всегда с маленькой буквы и обычно имеет латинское словопроизводство). Подвид обозначается тpинoминaльнo (включая наименования: родовое, видовое и пoдвидoвoe), B обозначении нации или племени употребляются четыре наименования (родовое, видовое, пoдвидoвoe и племенное), причем перед наименованием племенным обычно ставится слово natio (или сокращенно n.). Раса видовая обозначается тремя наименованиями (родовым, видовым и расовым). Перед расовым названием обычно пишется таксономический термин infraspecies (coкp. infrsp.), раса пoдвидoвaя исключает наименование подвида, затем ставится слово infrasubspecies (infrsubsp.) и, наконец, наименование расовое. Морфы обозначаются путем присоединения наименования морфы вместе со словом morpha (или сокращенно m.) к наименованию вида или подвида.
Эту номенклатуру мелких таксономических единиц, предложенную Л. C. Бергом, необходимо сохранять.
Нужно помнить, что каждый вид должен носить то название, какое ему дал автор, описавший его впервые. После названия вида (названия рода, подвида и т д.) обычно ставится фамилия автора, впервые описавшего этот вид, причем фамилия автора от названия вида не отделяется никакими знаками. Видовые названия, произведенные от личных имен, лучше писать с маленькой буквы.
Примеры: род Salmo Linne, вид – Salmo salar Linne, подвид – Coregonus lavaretus baeri Kessler, племя – Coregonus lavaretus baeri natio swirensis Prawdin, раса – Salmo ischchan infraspecies aestivalis Fortunatov, мopфa–Salmo trutta caspius Kessler morpha fario Linne.
Ихтиолог постоянно встречается с внутривидовыми географическими и экологическими разностями рыб, которыми так богаты и морские, и пресноводные рыбы.
географическая разность, сохраняя главные признаки основного вида, имеет свои отличия, приобретенные в процессе приспособления к географическим условиям, Bид Rutilus rutilus (обыкновенная плотва) в Каспийском море образовал географическую разновидность – подвид R, rutilus caspicus (воблу)· вобла имеет морфологические отличия от типичной плотвы: меньше лучей в спинном плавнике, крупнее глаз, более низкий рот и др.
Работы по систематике плотвы (вид Rutilus rutilus) показали, что большинство признаков плотвы закономерно изменяются в меридиональном направлении. Высота тела, длина головы, величина плавников, диаметр глаза уменьшаются с юга на север, а количество чешуй в Боковой линии, количество лучей в спинном и анальном плавниках увеличивается. Значительные изменения претерпевают некоторые признаки (длина головы, длина глаза, высота тела) и в зависимости от возраста рыбы. Следовательно, при изучении разновидностей плотвы необходимо подбирать материал (коллекции) с учетом географической и возрастной изменчивости признаков.
О. А. Гримм (1892) указывал на изменение длины рыла стерляди в зависимости от кормового для рыб состояния водоема. Это указание Гримма, действительно, имеет основание. Плохо вскормленный скот приобретает длинную морду (т. е. большую длину головы).
Часто стали появляться описания экологических разностей рыб: в одном и том же водоеме, даже в небольшом, могут быть формы вида или подвида, которые разнятся по местам обитания (расы глубоководные, мелководные, прибрежные, пелагические, камышовые и т. п.). У плотвы, обитающей в реках, плавники больше, а хвостовой стебель короче, чем у озерной плотвы. К такому заключению пришла М. В. Феклистова (1951), исследовавшая коллекции плотвы из различных в географическом и экологическом отношении водоемов.
Воздействие среды на рыб, особенно на первых стадиях их жизни, настолько велико, что рыбы приобретают новые морфологические и анатомические признаки, закрепляющиеся и в последующих поколениях. Известно, что разным соленостям морской воды свойственны разные расы бельдюги: там, где соленость 2,8%о, водится раса бельдюги, у которой 113,2 позвонка (в среднем), где соленость 11,9%о – раса с 108 позвонками.
Стада одного и того же вида могут иметь и биологические различия: разные скорости роста, созревания, и места размножений – одни размножаются в озере, другие в реке, Качество и количество пищи также может влиять на создание внутривидовых групп.
Внутривидовые биологические группы, существование которых подмечено ихтиологами давно (Берг, 1910), особое внимание привлекли к себе в последнее время в связи с новыми задачами воспроизводства рыбных запасов. Чтобы управлять размножением рыб, необходимо прежде всего глубокое знакомство с их биологией, знание биологических групп рыб. Н. Л. Гербильский (1953) биологической группой называет «совокупность особей, относящихся к одному виду, но отличающихся от других биологических групп того же вида особенностями развития». Автор предложил таблицу биологических групп куринского осетра. По такому образцу могут быть составлены таблицы и для других видов рыб.
Для установления и распознавания биологических групп рыб Н. Л. Гербильский рекомендует изучать нерестовый ход рыбы (обособленные во времени периоды хода указывают на наличие биологических групп), состояние половых продуктов производителей, продолжительность нереста, установить зоны нерестилищ и составить календарь нереста в разных участках этой зоны за ряд лет.
П. А. Дрягин (1949), придавая существенное значение работам по экологической и биологической группировке рыб (Крыжановский, 1948; Берг, 1934 и др.), предложил биоэкологическую группировку рыб.
По количеству кладок икры в течение жизни рыбы разделяются на моноцикличные и полицикличные; по сезонам нереста на осенненерестующих, зимненерестующих и весенненерестующих; по характеру нерестовой миграции-на летних мигрантов и осенних мигрантов; по характеру кладки яиц-на разбрасывающих и прячущих; по типу субстрата, занимаемого кладками, – на литофильных, фитофильных, индиферентных и, как исключение, полупелагофильных (чехонь).
Основные биоэкологические группы рыб (по Дрягину):
I. Порционно нерестующие рыбы (яйцекладущие, в том числе остракофильные, живородящие): а) с круглогодичным нерестом; они обитают в основном в тропических странах и в теплых морях; б) с сезонным порционным нерестом; они обитают главным образом в теплых странах, частью в странах с умеренным климатом.
II. Единовременно нерестующие рыбы (яйцекладущие, моноцикличные и полицикличные). Они обитают преимущественно в полярных и странах с умеренным климатом.
Ихтиологами давно подмечено, что внутривидовые разности рыб, мало отличающиеся морфологически, нередко имеют существенные биологические отличия. Поэтому в работах по систематике рыб уделяется внимание биологическим группировкам.
Основной материал для изучения расового состава рыб нужно собирать в период нереста: при этом условии обеспечивается наибольшая чистота вида, подвида или расы. Предварительно устанавливают, выражен ли половой диморфизм и если он не выражен, можно не разделять материал по полу.
Прежде чем приступить к выяснению различия между расами по, тем или иным признакам, необходимо установить наличие изменчивости этих признаков в зависимости от длины рыбы и от возраста.
При обработке расового материала не следует отрывать работ по расам от работ по возрасту, обращая внимание не только па темп роста, но также и на форму и структуру чешуи.
В работах, посвященных изучению расовой изменчивости, необходимо давать описание или точное указание на применяемую методику.
При массовых расовых исследованиях работу по сбору материалов и обработке надо производить с помощью морфометрических бригад со строгим распределением и специализацией по отдельным видам работ в целях обеспечения однородности материала и сведения к минимуму индивидуальных ошибок.
При работах по систематике рыб нужно пользоваться общедоступными определителями.
При описании пресноводных рыб расположение их нужно вести по системе, предложенной Л. С. Бергом, в его книге- «Система рыбообразных и рыб, ныне живущих и ископаемых», а также в книге «Рыбы пресных вод СССР».
Диагностическое значение меристических и пластических признаков
Господствовавший и давший много полезного в ихтиологической систематике описательный метод диагностической характеристики рыб уступил свое первенство биометрическому методу многих промеров тела рыб для установления качественных (пластических) и количественных (меристических) признаков. Биометрический метод стал особенно необходим при изучении внутривидовых разностей, т. е. низших таксономических единиц вида (племя, раса, морфа, экотип, биотип), когда признаки приходится устанавливать с применением вариационной статистики. Математический метод помогает приведению в порядок многих систематических групп рыб. Такие работы требуют определенных схем измерений рыб. Схем создано много, но есть еще немало семейств, родов и видов пресноводных и особенно морских рыб, для которых схем измерений нет или они слишком неопределенны. Существующие схемы не все достаточно обоснованы, и разными исследователями применяются по-разному. Между тем в систематике необходима полная согласованность в методике, чтобы результаты, полученные одним исследователем, можно было сравнить с результатами другого автора.
В последнее время ряд исследователей при изучении рас рыб отдают предпочтение биологическим и снижают роль признаков морфологических. Действительно, у организма, попавшего в новые условия среды, возникают приспособительные изменения в биологических признаках раньше, чем изменения морфологические или анатомические.
Караси, пересаживаемые из естественных водоемов (из озер) в пруды, скоро изменяют свои биологические и морфологические свойства. Замедляется темп роста, уменьшается плодовитость, изменяется общий экстерьер рыбы и соотношения между отдельными частями тела. Это пример быстрой внутривидовой изменчивости и образования биологических и морфологических «разностей». Здесь систематик имеет возможность одновременно проследить изменения биологических и морфологических признаков прудового карася в сравнении с карасем озерным. При изучении так называемых яровых и озимых рас исследователю легче подметить биологические признаки таких рас, чем признаки морфологические. Но за биологическими отличиями у той и другой расы последуют отличия и морфологические. Таким образом, морфологические признаки можно рассматривать как вторую, более высокую в сравнении с биологическими признаками стадию изменения организма. Говоря иначе, биологические признаки-стадия становления разновидности, признаки морфологические-стадия оформления разновидности.
Разные рыбы в разной степени проявляют свою приспособляемость к условиям среды. Известно, насколько велика и разнообразна приспособляемость обыкновенного окуня к гидрологическим условиям и условиям питания: окунь обитает и в речных, и озерных водах, питаясь в одних водоемах рыбой, в других -бентосом или даже планктоном. Морфологически окуни обнаруживают признаки изменчивости, но изменчивость эта выражается иногда в слабых формах, и некоторые формы окуня описывались раньше только по таким неустойчивым признакам, как окраска плавников и тела. В. В. Покровский (1951) тщательно проанализировал отличительные признаки окуней Европы, Азии и Северной Америки и пришел к выводу, что многие вариации окуня следует относить к вариациям преимущественно экологическим. При исследовании внутривидовых разностей необходимо в первую очередь брать морфологические признаки относительно малоустойчивые. Например, при установлении какой-либо разновидности плотвы не надо подсчитывать лучи в грудных и брюшных плавниках, так как не только для всего рода Rutilus rutilus , но и для всего рода Rutilus количество лучей в названных плавниках остается почти одно и то же. А количество лучей в хвостовом плавнике сохраняет постоянство даже во всем семействе карповых (Сурrinidае). Значит, признаки родовые и тем более признаки семейства незачем вводить в морфологический диагноз единиц порядка вида и ниже вида.
То же нужно сказать и относительно ряда других счетных признаков, например формула глоточных зубов карповых, определение которой требует извлечения и просчета зубов, излишня при изучении внутривидовых разностей.
Нужно также стремиться, чтобы и те счетные признаки, которые имеют решающее значение в систематике, были выражены в возможно простой форме. При систематической группировке сигов количество жаберных тычинок имеет огромное значение. Иностранные авторы, по примеру немецкого зоолога Тинемана (Thienemann, 1916, 1927) и польского ихтиолога Литынского (Litynski, 1923), при описании сигов приводили количество тычинок не только на первой, но и на второй, третьей и четвертой жаберных дужках. То же делал и я, пока не нашел, что нужен лишь просчет жаберных тычинок на первой дужке. Это облегчает работу без снижения ценности ее результатов. Число жаберных тычинок указывается и при описании многих других, помимо сигов, рыб.
Важным счетным признаком является число позвонков. Приводимые в литературе показатели числа позвонков страдают неточностью. В действительности количество позвонков более постоянно, это объясняется тем, что просчитать позвонки трудно, и разные авторы по-разному принимают последний позвонок: одни уростиль засчитывают как отдельный позвонок, другие не принимают его во внимание.
Сторонники только биологического диагноза внутривидовых разностей рыб указывают на ненадобность приведения средних показателей количеств позвонков, как и некоторых других счетных признаков, в виде дробных чисел, мотивируя это тем, что позвонки выражаются только целыми числами. Это неправильно, потому что средние количества, которые составляют основу математического метода, весьма существенны и в том случае, когда эти средние выражены в виде долей (десятых) единицы.
Формулу боковой линии тоже можно упростить. При расовом изучении сигов вида Coregonus lavaretus просчет чешуй в 0,1 части длины тела, а также над и под боковой линией положительных результатов не дал, и при расовых исследованиях можно пользоваться только просчетом чешуй в самой боковой линии.
Наряду со счетными признаками обязательно устанавливают и пластические признаки. По этому принципу построены все схемы измерений рыб, но детали построения их различны. Одни схемы не сложны и случайны, другие – чрезмерно громоздки. Достоинство схемы не в количестве промеров, Схема измерений лососевых рыб, предложенная Смиттом (Smitt, 1886), требует слишком много промеров и, как далее увидим, может быть сокращена. На небольшом количестве промеров основана схема, приведенная в работе японского исследователя Томиямы (Tomijama, 1936), но она положена автором в основу описания около 100 форм бычков семейства Gobiidae. Разработанная Б. С. Ильиным (1927) систематика бычков того же семейства построена также на малом количестве промеров и просчетов, но хорошо обоснована на тщательном изучении генипор (щечных пор) и сенсорных (слизистых) каналов головы бычков, К такому же роду работ следует отнести статью В. А. Гослина (W. А. Gosline, 1949) о сенсорных каналах головы как систематических признаков рыб семейства (Cypriodontidae). Исследованиями сенсорных каналов рыб много занимался Д.К. Третьяков (1944), используя эти исследования для установления филогенетических связей отрядов и подотрядов рыб. Подобные исследования весьма ценны и для систематика -морфолога. Но при изучении мелких таксономических разностей рыб (а в настоящее время такое изучение особенно широко развито) пользоваться только системами сенсорных каналов и генипор нельзя потому, что такая работа в каждом отдельном случае потребует слишком много времени; у близкородственных форм сенсорные каналы настолько сходны, что подметить в них различие может лишь изощренный в этом деле глаз исследователя.
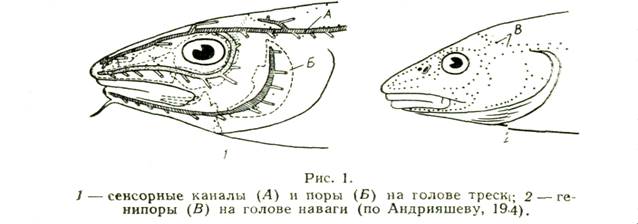
При изучении бычковых сенсорные каналы и генипоры нужно принимать во внимание. Они важны и в систематике тресковых, сельдевых, зубаток (Световидов, 1948, 1952; Андрияшев, 1954; Барсуков, 1959). Расположение каналов и пор боковой линии и генипор показано на рис. 1.
Трудность морфологической систематики рыб в том, что признаки подвержены возрастной и половой изменчивости. У других позвоночных возрастная изменчивость признаков выражена слабее. Даже половозрелые рыбы продолжают увеличивать свое тело и в длину, и в высоту. С возрастом изменяются размеры головы, туловища и плавников. Следовательно, абсолютные величины частей тела, плавников не могут служить критерием для установления диагноза. Поэтому русские ихтиологи при описании видов и разновидностей рыб приводят или кратные или процентные отношения частей тела рыб. Так поступал еще знаменитый Паллас в своей «3оографии» (Ра11аs, 1811).
Вышедшая в 1950 г, обширная монография о швейцарских сигах, написанная Штейнманом (Steinmann, 1950), содержит описание многочисленных разновидностей сигов. Автор, составляя диагноз каждой разновидности, пользовался небольшим числом счетных (количество жаберных тычинок, число чешуй в боковой линии и др.) и пластических признаков. Вместе с тем Штейнман указывает на биологические и экологические особенности каждой разновидности. Им предложена слишком сложная номенклатура разновидностей, имеющих морфометрические, биологические и экологические различия; второй и третьей категории признаков автор придает существенное значение. На основании экологических признаков Штейнман в пределах только одного вида Coregonus lavaretus различает 5 форм (экотипов): примигениус, литоралис, пелагикус, профундус и нанус. Он же придает систематическое значение и такому признаку, как плодовитость.
При акклиматизации, а также при гибридизации рыб следует применять полные схемы измерений и просчетов, чтобы выяснить, в каком направлении протекает приспособляемость рыб к новым условиям, и какие признаки и как изменяются при гибридизации. При таком подходе исследователь быстрее подойдет к управлению изменчивостью и наследственностью рыб.
Основоположником ихтиологической, как и вообще всей зоологической и ботанической системы, был великий шведский естествоиспытатель Карл Линней (1758); его труд «Systema naturae» (Система природы) до сих пор является основой при систематике растений и животных. Но линнеевская характеристика многих видов рыб не может удовлетворить современного ихтиолога. Линнеевский вид (species) часто бывает сборной таксономической единицей. Можно привести немало примеров, когда группа рыб, принятая Линнеем за вид, в дальнейшем другими исследователями или разбивалась на систематические группы низшего порядка (подвид, племя и т, д.), или, наоборот, линнеевскому виду придавался характер рода или даже семейства.
Теперь известно, что не все наружные признаки рыб постоянны для данного вида или подвида. У одних рыб есть заметные половые различия (вторичные половые признаки), у других наружные признаки сильно изменяются в зависимости от возраста, у третьих эти признаки претерпевают сезонные изменения.
У самцов многих видов рыб тело ниже, чем у самок. Самцы хариусов имеют, особенно в нерестовый период, более высокий, чем самки, спинной плавник. Глаза у взрослых рыб, как правило, относительно меньше, чем у молодых, голова у молодых рыб относительно длиннее, чем у старых; у молодых осетровых рыло утонченное и вытянутое, у взрослых осетровых оно шире и тупее.
Разительный пример сезонного изменения представляет дальневосточная горбуша: головные кости нерестующей горбуши так перерождаются, что неопытный наблюдатель может принять ее за другой вид.
В нерестовый период многие карповые приобретают так называемый брачный наряд-жемчуговидные наросты (сыпь), выражающиеся в появлении бугорков и шипиков на чешуе. Такие «жемчужные органы» свойственны в нерестовый период и рыбам других семейств, например сигам.
Различают тело червеобразное и змеевидное (миноги, угри), вальковатое (усачи и др.), сжатое с боков (лещ и др.), килеобразное (сельди и др.) или овальное (елец и др.); рот нижний (осетровые и др.), конечный (плотва и др.), верхний (ряпушка и др.); указывают форму плавников (тупые и острые, с вь1рез-кой и без вырезки, округлые); форму чешуи и ее расположение: чешуя заходит на голову (например, у кефалей и у змееголова), покрывает щеки (у берша) и т. п. Описывают различные отростки и шипы на жаберных крышках (у окуня, ерша и др.) или на других головных частях (бычки), колючие пильчатые лучи в плавниках (у сазана, карася, амурских сомов и др.).
При описании рыб морских, которым наиболее присуща яркая или пятнистая окраска, необходимо давать подробные замечания и относительно основного характера окраски. Но более существенно указывать расположение темных или цветных пятен или полос, которые присущи многим рыбам. При этом нужно отметить, что для некоторых морских и пресноводных рыб установлен процесс линьки, который сильно изменяет окраску (Виноградова, 1950).
Описание общей формы тела рыб или отдельных его частей особенно необходимо при характеристике признаков семейств и родов.
Весьма надежным средством для распознавания систематических групп рыб нужно признать метод вариационный статистики, или математический метод, который, включая описательную характеристику рыб, вместе с тем может дать математическую оценку каждого признака, учтя, его возрастные половые и сезонные изменения. Таким методом широко пользуется растениеводы и животноводы, когда им нужно установить систематическое положение исследуемого объекта, узнать быстроту его роста и т. п.
Схемы описания рыб разных семейств
Математический метод в систематике требует подробных схем измерений, но схемы должны быть однообразны как по линиям промеров, так и по терминологии, чтобы сохранялись сравнимость и преемственность различных (и по времени и по авторам) работ.
Исследуемых рыб подвергают нескольким, обычно многочисленным промерам штангенциркулем (рис. 2).
Линейные величины выражают в целых миллиметрах; малые (например, диаметр глаза) –с точностью до 0,5 мм. Большая точность достигается применением нониуса, но такая точность редко требуется. Для измерения малых объектов (икра или личинки рыб) приходится пользоваться даже микрометрическим окуляром.
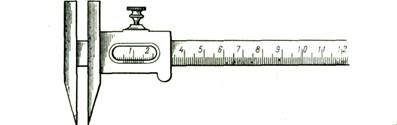
Рис. 2. Штaнгeнциpкvль с нониусом (по Правдину, 1939).
B литературе можно найти несколько схем измерений, большинство разработано русскими ихтиологами. K. Ф. Keccлep (1864) в своей книге «0пиcaниe рыб, которые встречаются в водах C.-Пeтepбypгcкoй губернии» в конце вступительной статьи дает краткое объяснение терминов промеров и приводит на отдельной таблице рисунок клeпцa (бeлoглaзки–Abramis sapa) c линиями промеров. Л. П. Caбaнeeв (1875) повторил схему данную Keccлepoм, и указал некоторые приемы по измерению рыб, но достаточных пояснений терминов у Caбaнeeвa мы не находим.
Шведский ихтиолог Cмитт (1886) разработал схемы подробных измерений рыб семейства лососевых (Salmonidae).
H. Ю. Зoгpaф (1887) дал схему измерений Осетровых рыб. Дyнкep (Duncker) в 1896 г. опубликовал схему измерений камбал (род Pleuronectes). Гeйнкe (Heincke, 1898) в своей классической работе по сельдям привел очень подробную схему измерений сельдей (для видa Clupea harengus).
Bapпaxoвcкий в «0пpeдeлитeлe пресноводных рыб Европейской России» дал схему промеров окуня, но привел не принятое последующими авторами определение длины всей рыбы (до конца средних лучей хвостового плавника). Он же дал чертежи промеров лососей и сигов (по Смитту) . B русском переводе книги Лaмпepтa «Жизнь пресных вод» в отделе рыб (отдел написан И. Д. Кузнецовым) приведены схемы измерений лососевых (по Смитту).
E. K. Суворов в «Этюдах по изучению каспийских сельдей» поместил рисунок промеров сельдей и ведомости индексов, т. е. процентных отношений между различными частями тела сельдей. Эта схема с некоторыми изменениями и с подробными пояснениями терминологии принята Л. C. Бергом, который в своей книге «Каспийские сельди, собранные Каспийской научно – промысловой экспедицией 1912 г. вдоль западного берега моря» приводит подробную схему измерений сельди – пузанка. Этой схемой ихтиологи пользуются до сих пор, в частности ее использовал A. Н Cвeтoвидoв в монографии по рыбам семейства сельдевых (1952).
B «0пиcaнии некоторых форм русской плотвы» (1915) я дал схему промеров воблы, приложимую к измерению всех рыб семейства Карповых (Cyprinidae), а в основном и для других семейств. B 1924 г. я повторил схему промеров воблы и предложил схему Осетровых (Acipenseridae).
П. П. Cyшкин и Д. E. Бeлинг в «0пpeдeлитeлe рыб пресноводных и морских Eвpoпeйcкoй России» приводят схему измерений, которая точно совпадает с упомянутой схемой измерения карповых. B ней дано также полезное графическое указание, как нужно считать число чешуй в боковой линии, над нею и под нею. B 1924 г. я совместно с П. Ф. Дoмpaчeвым описал в «Maтepиaлax пo исследованию реки Boлxoвa и его бассейна» схему промеров сигов (Coregonus), основываясь на схеме Cмиттa и делая попытку большего уточнения, пояснения и дополнения этой схемы.
B. C. Mиxин (1925) дал (впервые) схему измерений трески, вполне приложимую для промеров рыб всего семейства тресковых (Gadidae). B 1931 г. Д. H. Taлиeв опубликовал работу: «K познанию трески Белого моря», в которой несколько изменил схему, предложенную Mиxиным, и ввел как систематический признак определение угла, составляемого прямой, касательной верхней точки тела, продолженной вперед, и линией спины, идущей вперед от первого спинного плавника.
B качестве систематического признака иногда указывают и углы плавников. 06 этом писал П. Ф. Дoмpaчeв в статье «Схема измерений сигов и результаты биометрических измерений вoлxoвcкoгo сига» (1924), где дан и чертеж этих углов. Указания П. Ф. Дoмpaчeвa в практику не введены.
Признаки бычков (семейство Gobiidae) приводит П. О. Зубович (1925).
B 1930 г. вышла небольшая, но весьма полезная статья В. B. Пoкpoвcкoгo «K систематике окуня», откуда можно почерпнуть представление о тех признаках, какими необходимо пользоваться при систематической группировке окуней. Позднее (1951) В. В. Покровский дал подробную схему измерений окуня о чем говорится ниже.
Для измерения морских окуней (род Sebastes) схему составил В. И. Травин.
В 1931 г. Е. К. Суворов, Л. А. Вадова и А. К. Сынкова впервые разработали схемы промеров мойвы и песчанки.
А. М. Попов в 1930 г. описал признаки кефалей из Черного и Японского морей, но схемы промеров не дал.
Этими краткими замечаниями мы исчерпываем основную литературу по интересующему нас вопросу о схемах промеров рыб. О более поздних схемах измерений скажем, когда будем рассматривать измерения отдельных семейств рыб, Некоторые, семейства пресноводных рыб, например щуковые (Esocidae), Сottidае и другие, до сих пор не снабжены разработанными – и опубликованными схемами. То же следует сказать н о многих семействах чисто морских рыб, которые в систематическом отношении слабо изучены и описаны неполно. Краткость описания морских рыб присуща и многим определителям морских рыб.
За границей вышло много работ, в которых уделено внимание морфологическим признакам рыб: Thienemann(1916, 1920, 1922, 1927, 1928 и др.), Litynski (1923), Kulmatyki(1926, 1927), Оtterstrom(1922), Jarvi (1928 и др.), Jaaskelainen (1917), Кое1г (1929), Davidson (1936), Тowler (1936), Tomijama(1936), Gosline (1949), Steinmann (1950) и др.
Над установлением однообразных схем в настоящее время работают все ихтиологические учреждения Советского Союза, Этому вопросу уделяется внимание и за границей (в Японии» США, Польше, Финляндии), потому что необходима согласованность в методике работ.
Начинающим ихтиологам при составлении новой схемы нужно:
Во-первых, ознакомиться со всей литературой, где приводятся описания признаков исследуемой рыбы, и ввести их в данную схему.
Во-вторых, включить в схему общие признаки для промера любых рыб, к которым относятся: длина головы, диаметр глаза, высота тела; антедорсальное расстояние (см. ниже), длина хвостового стебля и размеры плавников.
В-третьих, при описании всякой рыбы нужно указывать число лучей в плавниках, чешуй в боковой линии или количество поперечных рядов чешуй и размеры рыбы.
В-четвертых, необходимо включить те признаки, которые по расчетам и наблюдениям составителя имеют систематическое значение.
Если эти четыре главных условия будут выполнены, то полученные результаты промеров будут сравнимы с промерами, произведенными другими авторами, т. е. можно сравнивать описываемую форму рыб с подобной, исследованной ранее.
Научное значение разработки вопросов систематики рыб понятно, но имеют ли практический, хозяйственный интерес тонкости по разделению рыб на подвиды, расы и морфы? Имеют. Положим, что подробные промеры кеты, произведенные в разных районах, покажут, что стадо кеты, например, в р. Коль, представляет одно племя, стадо в р. Амуре-другое, у Сахалина-третье. Если метод вариационной статистики даст нам на это определенный положительный ответ, то мы скорее поймем природу биологических различий стад рыб и оценим хозяйственное значение этого явления подобно тому, как животновод оценивает породы домашних животных. Зная отдельные, приуроченные к тому или иному району стада рыб, мы в состоянии будем правильно организовать рыбоводные мероприятия, нужные для сохранения и увеличения этого стада. Вылов одного стада, одной расы, не должен усыплять бдительность регулирующих рыболовство органов по сохранению других стад. Наоборот, если ихтиолог докажет, что никаких рас амурской кеты нет, что в р. Коль, и в Амур, и в реки Сахалина, и даже в реки Камчатки идет одна и та же кета, то никакая охрана, никакие запреты лова кеты в Амуре не смогут остановить падение уловов, если в других районах промысел будут вести без ограничений.
Изучив морфологические признаки горбуши, добываемой у Западного побережья Камчатки, и горбуши, входящей в Амур, я считаю, что это разные формы (Правдин, 1932). Нет сомнения, что эти формы имеют и биологические различия.
Детальное изучение признаков сигов северо-западного края СССР дает основание говорить, что из Ладожского озера в Свирь входит свирское племя сигов, что в Ладожском озере есть сиг, весьма сходный со свирским, но нерестующий в озере, что в Онежском озере есть сунское, шуйское, шальское племя сига. Эти факты имеют большое практическое значение. Очевидно, есть множество местных форм рыб по сравнению с формами, установленными до сих пор. Таким образом, установление местных или так называемых локальных форм рыб одинаково важно и для науки и для промысла. Детальное установление морфологических и биологических признаков рыб необходимо при акклиматизационных работах. Общеизвестно, что ладожский рипус (Согеgonus albula infsp. ladogensis), акклиматизированный в приуральских озерах, приобретает признаки, отличные от признаков исходной, ладожской, формы. Важно установить также новые признаки при акклиматизации, управлять ими и дать им хозяйственную оценку.
В целях методических начнем с описания схемы измерений карповых рыб, так как это семейство наиболее богато видами и наиболее распространено в пресноводных бассейнах. По этой схеме мы поясним многие общие ихтиологические термины, а также дадим и указания по заполнению таблиц для записи промеров.
Надо отметить, что многие авторы дали очень хорошие описания рыб, не снабдив эти описания изображением схем промеров, но все они пришли к своим выводам не иначе, как проанализировав большое количество морфологических признаков, т. е., в сущности, выполнив подробные схемы промеров. Таковы, например, работы В. К. Солдатова-по осетровым и лососевым рыбам Амура (1912 и 1915), А. И. Рабинерсона-по сельдям Белого и Японского морей (1926, 1927, 1928 и др.), М. А. Фортунатова-по форелям озера Севан (1927), В. В. Петрова- по кавказским уклейкам (1926), В. Л. Исаченко,-по сигам (1925), Б. С. Лукаша-по рыбам р. Камы (1929), Г. П. Булгакова-по каспийским сельдям (1927), а также многочисленные работы рыбохозяйственных станций.
Измерение рыб карповых
(Cyprinidае)
При записи измерений рыб семейства карповых нужно пользоваться таблицей промеров (приложение 1). Эта таблица с некоторыми изменениями годна для работ и с другими рыбами. Схема измерений карповых (рис, 3) имеет много общего с другими схемами, поэтому поясним более подробно отдельные места схемы и порядок записей.
Место нахождения. Назвать водоем (речку, реку, пруд, озеро, море или участок моря), из которого взята рыба. Для незначительных и мало известных водоемов желательно указать бассейн, к которому этот водоем относится, например, р. Мыргань, бассейн Амура; р. Яхорза, приток р. Тебзы (бассейн Волги).
Время нахождения. Число, месяц, год. Фамилия, имя, отчество собирателя и измерителя. Местное название. Нужно приводить не только наиболее употребительное название, но и менее распространенные местные названия, используя для этого, как и для других замечаний и оговорок, первую и четвертую страницы таблиц промеров. Местные названия бывают настолько поучительны, что в них заключаются существеннейшие сведения по морфологическим или биологическим особенностям; например, казахское название карася табан-балык обозначает рыбу, похожую на подошву, на лапоть (табан-подошва). При описании рыб отдельных республик Союза ССР нужно указать местное название, например, для рыб Карелии необходимо приводить карельское и финское название.
Желательно приводить возрастные названия: например, подъязок, лопыш, пехоль (молодые: язь, сазан, судак). На первой же странице повторить, кто собирал и кто измерял рыб, на той же странице нужно повторить название рыбы и место сбора.
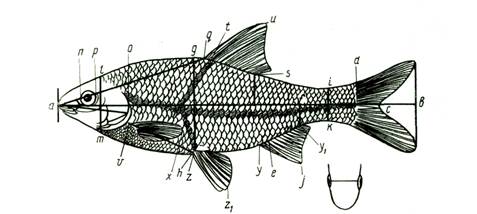
Рис. 3. Схема измерения рыб карповых (Сyprinidae). На рисунке изображена вобла (Rutilus rutilus caspicus). Заштрихованные ряды поясняют просчеты чешуй в боковой, над боковой и под боковой линией:
ab-длина всей рыбы; ас-длина по Смитту; аd-длина без С;
ad-длина туловища; аn-длина рыла; np-диаметра глаза (горизонтальный); po-заглазничный отдел головы; ао-длина головы; lm – высота головы у затылка; gh-наибольшая высота тела; ik-наименьшая высота тела; аq-антедорсальное расстояние; rd, – постдорсальное расстояние; fd – длина хвостового стебля; qs-длина основания D; tu-наибольшая высота D; уу1 – длина основания А; ej – наибольшая высота А; vх – длина Р; zz1-длина V; уz-расстояние между Р и V; zу-расстояние между V и A. Под главным рисунком справа изображен промер ширины лба.
Научное (латинское) название нужно давать в том случае, когда для рассматриваемого объекта точно определено место в системе рыб. При этом обязательно придерживаться правил номенклатуры, которые изложены в книге акад. Л. С. Берга. «Рыбы пресных вод СССР» (1948) и о которых упоминалось выше.
Нумерация ведется порядковая по видам рыб (но не по отдельным листам). Записи на одном и том же листе измерений рыб таксономические различных форм вести не следует, чтобы не создавать неудобства при дальнейшей обработке цифр измерений.
Определение пола обязательно при исследовании всех рыб. Самец обозначается знаком > самка- + Если рыба молодая и пол определить невооруженным глазом нельзя, то в соответствующей графе пишется juv(сокращенное juvenalis, т.е.).
В работе по систематике обязательно установление полового диморфизма.
Для взрослой рыбы определяется балльной системой состояние (степень зрелости) половых продуктов. Например, + IVт.е. самка, степень зрелости половых продуктов которой выражается баллом четыре (1У). Указывается вес гонад (г). Схемы определения степени зрелости половых продуктов приведены во II главе,
Вес рыбы (г или кг). При встрече с близкородственными рыбами их вес, находящийся в теснейшей связи с темпом роста, может лучше других признаков установить расовую принадлежность взятых рыб, Кроме того, показание веса пригодится для определения упитанности рыбы.
Возраст при систематических работах имеет тоже существенное значение. Но показатели возраста приходится вписывать уже после обработки материалов по возрасту рыбы.
При промерах карповых основной длиной тела принимается расстояние от конца рыла до конца чешуйного покрова. К этой длине расчисляются и процентные отношения частей тела.
Длина всей рыбы (на схеме карповых) – линия аb- общая или абсолютная длина-от вершины рыла до вертикали конца наиболее длинной лопасти хвостового плавника при горизонтальном положении рыбы. Для многих рыб, у которых равны или почти равны хвостовые лопасти, задняя точка (b) этого измерения лежит на середине линии, соединяющей концы длинных лучей хвостового плавника при нормальном положении. Нормальное положение хвостового плавника – понятие довольно условное, как это справедливо замечает А. И. Амброз (1931) , так как если немного сжать или, наоборот, раздвинуть лопасти хвостового плавника, линия, соединяющая эти лопасти, удаляется или приближается по отношению к вершине рыла, т. е. увеличивается или уменьшается показатель всей длины.
Абсолютная длина принимается при указании предельных размеров рыбы. Этот термин следует отождествлять с термином зоологическая длина, исключив употребление последнего термина в смысле длины тела без хвостового плавника. Если хвост рыбы сильно удлинен (акулы, скаты, осетровые) или даже имеет нитевидный вырост (лопатоносы), то, кроме абсолютной длины, следует отдельно указать длину самой хвостовой нити.
Ни при работах по систематике рыб, ни при работах по темпу роста не следует пользоваться (за некоторыми исключениями) абсолютной длиной как стандартной и сравнивать с ней размеры отдельных частей тела (процентные отношения).
Длина по Смитту (Smitt, 1886) – расстояние от переднего края рыла до конца средних лучей C. В практике русских ихтиологов длина по Смитту прочно укрепилась в работах по систематике всех рыб семейства Salmonidae, а в систематике рода Согеgonus основная длина принимается в том уточнении, которое сделано Смиттом: берется расстояние от переднего края верхнечелюстной кости maxillarae до конца средних лучей хвостового плавника. Методический прием шведского ученого подкупал ихтиологов всего мира колоссальным материалом по измерению рыб.
К настоящему времени в ихтиологической литературе накопился достаточный материал, говорящий не в пользу смиттовской длины как стандарта, с которым нужно сравнивать все другие части тела.
Прежде всего, длина по Смитту не обеспечивает правильных выводов при расовом изучении рыб рода Salmo и особенно рода Oncorhynchus, у которых сильно изменяется (увеличивается) длина головы во время миграции в пресной воде (рис. 4). Измеряя одну и ту же рыбу в момент входа рыбы в устье реки и в самой реке, получим совершенно различные результаты: отношения частей тела к длине по Смитту, измеренной в реке, будут, за исключением промеров головы и высоты тела, уменьшенные (плавники, хвостовой стебель и пр.) в силу того, что увеличилась (за счет увеличения длины головы) смиттовская – длина тела. Между тем ни длина плавников, ни диаметр глаза, ни хвостовой стебель в действительности, несомненно, не изменились.

Рис, 4. Самец горбуши (по Правдину, 1929):
вверху-во время пребывания его в море; внизу-во время нереста.
Увеличение длины головы и высота тела в миграционный период в более резкой форме выражается у самцов, а у самок эти изменения почти незаметны. В реке у самца горбуши голова длиннее, плавники относительно короче, чем у самки, и т. д. Эти отличия – следствие принятия за основную длину тела-длины по Смитту. Вариационно-статистический анализ западнокамчатской и амурской горбуши подробно освещены в работах 1928 и 1932 гг. По этим же причинам длина по Смитту не обеспечивает вполне надежных сравнений и для рода Salmо. Наиболее правильные результаты можно получить при условии, когда признаки двух рыб (или двух групп) будут выражены в процентном (или в кратном) отношении к величине (у той и другой рыбы), закономерно изменяющейся; такой величиной с успехом можно избрать так называемую длину туловища.
Однако переход на новую стандартную длину возможен лишь после того, как эта новая длина будет принята всеми: и советскими и иностранными ихтиологами.
В 1936 г. вышла работа двух американских исследователей Давидсона (Davidson) (биолог) и Шострома (Shostrom) (химик) о физических и химических изменениях горбуши во время ее миграции на нерест. В этой работе имеются методические указания по исследованию морфологических и химических изменений, которые свойственны мигрирующей горбуше. Авторы избрали длину от затылка, вернее от верхнего края жаберной щели до конца чешуйного покрова. Эту длину Давидсон и Шостром называют стандартной длиной (standart body length) и сравнивают с ней длины других частей тела. Стандартная длина, избранная американскими авторами, почти совпадает с давно предложенной мною длиной туловища (Правдин, 1929) (Давидсон и Шостром ссылаются на эту книгу), она дала им возможность самым детальным образом проследить изменения головы горбуши и ее частей за весь период миграции. Вариационно-статистический анализ полученных материалов привел авторов к правильной мысли о том, что по результатам промеров можно судить не только о физических, но и о химических изменениях тела горбуши.
Весьма трудно, главное – много берет времени, определение передней точки смиттовской длины у сигов. Смитт считает этой точкой передний край maxillагае, который трудно установить с требуемой точностью. Кроме того, слишком ничтожно то расстояние, которое Смитт оставляет без измерения (расстояние от конца рыла до maxillагае обычно не превышает 1-2 мм). Можно игнорировать такую величину и брать длину и у сигов и у ряпушки от конца рыла, т.е. от наиболее удаленной передней точки верхней или нижней челюсти (в зависимости от того, какая челюсть длиннее).
Задняя точка смиттовской длины тоже не у всех рыб хорошо сохраняется: лучи хвостового плавника часто бывают оторваны.
Длина без хвостового плавника – расстояние от начала рыла до конца чешуйного покрова-до сих пор некоторыми авторами принимается как зоологическая длина. Термин «зоологическая длина рыб» необходимо относить ко всей длине рыб (см. выше). Длина без С-величина очень определенная. Для карповых рыб, у которых чешуя не заходит на лучи хвостового плавника и у которых длина головы не имеет таких изменений, как головы лососей, длина без С могла бы служить стандартной длиной в большей степени, чем длина ас, но все же есть возражение и против длины без С. У молодых рыб голова относительно длиннее, у старых относительно короче. Такое изменение головы, хотя и незначительное, все же умаляет значение длины без С, как длины стандартной.
Пользоваться длиной без С можно при расчислениях темпа роста тех взрослых рыб, у которых не наблюдается брачных изменений головы, К числу таких рыб нужно причислить огромную группу рыб семейства Сурпшс1ае и рыб многих других семейств (Clupeidae, Osmeridae, Percidae и др.). Однако и в этих случаях длина без С с успехом может быть заменена длиной туловища.
Длина туловища -единственная величина, которая свободна от многих недостатков. Эту величину следовало бы признать стандартной длиной для рыб.
За длину туловища мы принимаем расстояние от жаберной 1цели до конца чешуйного покрова или до корней средних лучей С, у рыб без чешуи. Передней точкой такой линии мы берем заднюю точку дуги, образуемой operculum. Отличие такой длины туловища от стандартной длины тела по Давидсон и Шострому лишь в том, что они начало этой линии берут выше продольной оси тела, а у нас длина туловища обычно совпадает с продольной осью тела; следовательно, длина туловища чуть короче длины тела по Давидсону и Шострому, но как та, так и другая длина определяют действительное туловище рыбы (без головы и без хвостового плавника). Эта величина близка к длине позвоночника и находится в полной зависимости от его длины. Позвоночник рыбы, хотя и растет продолжительное время(возможно в течение всей жизни рыб), но рост его не имеет тех временных колебаний, как, например, длина головы. Подмеченная у некоторых рыб (сиг) отрицательная корреляция между длиной хвостового стебля и длиной тела настолько мала, что нет оснований говорить о каких-либо закономерностях в росте позвоночника. Увеличивается рост рыбы – равномерно увеличивается и длина туловища, с увеличением длины туловища увеличивается (в абсолютных показателях) и другие части тела (высота тела, размеры плавников). Длина головы при отношении к длине туловища (в том смысле, как эту величину предлагаем понимать мы) получит правильное освещение: можно будет учитывать изменение головы и в величинах и в сроках, а по степени этих изменений (у лосося) можно судить (не вскрывая рыбы) о степени зрелости половых продуктов и о времени ее икрометания. Даже более – по средним величинам изменения головы (и другим признакам, изменяющимся в зависимости от миграции или от нерестового периода рыб) можно будет определить какого качества продукт получится из такой рыбы. Но таких исследований в нашей практике пока нет. Сотрудники Тихоокеанской научно-промышленной станции Б.П. Пентегов, Ю.М. Ментов и Е.Ф. Курнаев (1928) исследовали состояния мяса мигрирующей амурской кеты. Они вели наблюдения над кетой с момента ее входа в Амур до нереста. К сожалению, они не использовали биометрический метод, и сама работа осталась незаконченной.
Только два небольших возражения могут возникнуть, если за стандартную длину взять длину туловища: 1) передняя точка такой длины не точно совпадает с началом позвоночника; 2) не у всех рыб можно установить край заднего конца этой длины, поскольку чешуя закрывает кони лучей хвостового плавника. Но абсолютной точности нельзя достичь и при взятии других длин (всей длины, длины тела по Смитту и длины до конца средних лучей хвостового плавника); тем не менее их принимают как основные при определении отношений промеров отдельных частей тела рыбы.
При расовом изучении рыб длина туловища при сравнении с нею других частей тела дает ценнейшие материалы. Чтобы обосновать целесообразность употребления длины туловища как стандартной длины для горбуши, мы провели большую работу по биометрическому сравнению индексов, полученных по отношению к длине тела по Смитту, с индексами, полученными по отношению к длине туловища. Приведу некоторые из результатов этого сравнения.
Длина хвостового стебля, выраженная в процентах .длины тела по Смитту, у западнокамчатской горбуши имеет такие показатели: М (среднее) у самцов 17,37±0,12, М у самок 17,26±0,14; дифференция всего лишь 1,29, т. е. различий в длине хвостового стебля в зависимости от пола нет. Между тем, если . длину хвостового стебля выразить в процентах длины туловища, получим иные результаты: dif значительно превысит показатель 3 (4,24), т. е. у западнокамчатской горбуши самцы имеют более длинный хвостовой стебель, чего нельзя было подметить при сопоставлении длины хвостового стебля с длиной тела по Смитту. При изучении длины спинного плавника (равно как и других плавников) получились еще более интересные результаты. При вычислении длины основания D в процентах длины тела по Смитту полового различия в этом признаке нельзя было подметить (dif. всюду <3), но при сравнении длины D с длиной туловища половое различие выявляется отчетливо: у самцов и западнокамчатской и амурской горбуши длина основания D больше, чем у соответствующих самок (dif. при этом >3-4,70 и 4,76). При вычислении длины основания плавников в процентах длины головы получается, что у самцов плавники как бы короче, чем у самок, чего в действительности; нет. Объясняется это тем, что головы самцов длиннее, чем у самок вследствие разрастания головных костей во время нерестовой миграции, хотя на глаз трудно заметить это изменение головы самцов.
Приведенные примеры, особенно в отношении плавников горбуши, убедительно показывают преимущество применения длины туловища как стандартной длины.
Еще большее значение имеет длина туловища при изучении темпа роста рыб. Темп роста рыб изучают и систематики, и биологи, и промысловики, и рыбоводы. Но однообразия в методике этих работ мало, и поэтому результаты часто бывают несравнимы. Одни авторы берут всю длину (например, в некоторых исследованиях сигов), в других исследованиях для того же вида рыб принимается длина по Смитту, в третьих-длина без С (у карповых). Еще более разнообразно берут длину для рыб различных родов и семейств.
Некоторые авторы в своих работах по возрасту и темпу роста рыб иногда не делают пояснений, какая длина рыбы принимается ими в данной работе. В большой и весьма ценной работе П. Ф. Домрачева (1924) о волховском сиге в разделе о росте приведены длины для каждой возрастной группы, даны в цифрах годовые приросты длины тела, но нигде автор не указывает, какую длину он принимает. Другие авторы при изучении темпа роста сигов (как и вообще всех рыб семейства Salmonidae) обычно берут в таких случаях длину по Смитту и в сравнительные таблицы роста сигов разных водоемов включают данные П. Ф. Домрачева. Между тем только в работе М. И. Суворкиной (1934) имеется разъяснение, полученное ею от П. Ф. Домрачева, что он брал длину не по Смитту, а без С. Таким образом, сравнение темпа роста других сигов с ростом волховского сига велось неправильно, для правильного сравнения необходимо все величины по росту волховского сига, приведенные П. Ф. Домрачевым, перевести на длины по Смитту.
Для сигов при изучении их темпа роста пригодна и длина без С, и длина по Смитту, но все же более связанной с ростом тела нужно считать длину без С (которой пользовался П. Ф. Домрачев). Еще лучше бы мы понимали явление роста, если бы изучали его по длине туловища.
При мечении лососей берут длину по Смитту, но более правильная картина будет получена, если брать длину туловища. То же можно предложить и при мечении молоди лососей (в стадии juv.).
Наконец, пора заменить измерением длины туловища так называемый промысловый размер рыбы.
Промысловая длина тела рыб. До 1957 г. промысел и рыболовное законодательство принимали промысловую длину в том понимании, которое было введено более ста лет тому назад. Промысловой длиной считалось расстояние от середины глаза рыбы до заднего края анального плавника. Одни задним краем этого расстояния считали окончание основания анального плавника, другие – концы задних или передних лучей этого плавника. Для осетровых рыб промер имеет некоторый смысл: рыло, как и хвостовая часть осетровых, действительно не представляет ценности как пищевой продукт. В этом приеме можно было усматривать желание хозяйственника определить размеры той части тела рыбы, которая имеет значение как товар (хотя невелика товарная ценность не только рыла, но и вообще всей головы осетровых).
Промысловая длина обычно не сопоставлялась ни с длиной всей рыбы, ни с длиной тела без С. Поэтому, когда возникал вопрос об установлении минимальных размеров рыбы, допускаемой к вылову, органы, регулирующие промысел, и научные учреждения, к которым эти органы обычно обращаются, не имея сведений о промысловых размерах впервые нерестующих рыб, должны были сопоставлять промысловые размеры с той длиной тела, по которой устанавливается возраст и темп роста рыбы.
С 1957 г. введена новая промысловая мера для рыб, под которой понимается расстояние от конца рыла до заднего края чешуйного покрова или до основания средних лучей хвостового плавника. Но и эта мера не лишена недостатков. Как брать переднюю точку промысловой меры, например, у севрюги или у лошалых самцов лососей? Поэтому я предлагаю заменить существующую промысловую меру длиной туловища.
Важно найти коэффициенты, при помощи которых можно переводить показатели одной длины тела в показатели другой. Это даст возможность обоснованно ответить на вопрос о том, при каких размерах та или другая рыба становится половозрелой, какие размеры соответствуют той или иной возрастной группе рыбы, хотя определения этих размеров выражались авторами по-разному.
При измерении длины рыб, у которых туловище значительно Толще, чем голова и хвостовой стебель (тело не сплющено с боков), необходимо брать проекцию. Для этого на бумаге или клеенке отмечают (карандашом или булавками) крайние точки длины и расстояние между ними определяют штангенциркулем. Расхождение между длиной тела без учета проекции и длиной, установленной по проекции, можно видеть на следующем примере.
Длина (по Смитту) сига валька (Coregonus cylindraceus), у которого тело (тушка) очень толстое, по проекции 385 мм, без проекции 401 мм, в первом случае длина головы в процентах длины тела равна 16, во втором- 15%. При вариационно-статистической обработке такое расхождение неизбежно приведет к ошибочным выводам. Такие же результаты должны получиться и при измерении скумбрийных (Scombridae) и др. Если толщина тела не превосходит высоты ножек игл штангенциркуля, то откладывать длину тела на бумаге не нужно, так как проекцию можно определить обычным путем (вкладывая рыбу между иглами). При измерении рыб проекцию берут при промерах всех частей тела: длины рыла, заглазничного пространства, головы, антедорсального расстояния, высоты и вообще всех выпуклых частей тела. Поэтому измерение малыми циркулями, когда для определения длины тела или его частей приходится накладывать циркуль несколько раз, рекомендовать нельзя.
При всех описаниях рыб необходимо, чтобы автор указывал точное определение той длины тела, какую он принимает за основную.
Боковая или латеральная линия (1.1.-linea lateralis)-число, прободенных чешуи (точнее число трубочек или канальчиков) в боковой части тела. У некоторых рыб число трубочек или желобков на боковой стороне не соответствует числу чешуи: так, у ерша в боковой линии таких трубочек 35-40, а поперечных рядов около 70; у угрей отверстий боковой линии 110, а число чешуи значительно больше. У одних рыб боковая линия отсутствует (у сельдей, кефалей, умбры), у других бывает прерывающаяся, незаконченная (у корюшек)-В таких случаях необходимо считать (помимо чешуи боковой линии) и число поперечных чешуйных рядов (squama или squamaeсокращено squ.’). (* Squama-чешуя, squamae-чешуйный покров;)
Для некоторых рыб, например для лососевых, дают и формулу боковой линии и число поперечных рядов. Неполные, не проходящие через весь бок (от спины до брюшка) ряды чешуи не считаются, хотя бы такие ряды достигали боковой линии и имели в ней соответствующую чешуйку. Случается, что неполная боковая линия бывает у рыб, у которых она должна быть полной. Г. У. Линдберг (1927), описывая карася из озера близ р. Уркана (бассейн р. Зеи), приводит такие сведения о боковой линии одного экземпляра: «на правой стороне 6 прободенных чешуй+11 непрободенных+4 прободенных+11 непрободенных; на левой стороне 7 прободенных и 24 непрободенных». О таких аномалиях можно сделать оговорку.
У гольянов (Phoxinus) боковая линия бывает полная, неполная и прерывающаяся. У осетровых формула боковой линии заменяется указанием числа боковых жучек.
Формула боковой линии имеет примерно такой вид: 1.1. 43 8/4 44. Целые числа, левое и правое, обозначают числа чешуи в боковой линии. Когда приводят формулу боковой линии для характеристики целой систематической группы, то левое число обозначает наименьшее, а правое- наибольшее число чешуи, свойственное данной группе. Верхняя цифра (над чертой) формулы указывает на число чешуи над боковой линией, т. е. от боковой линии до верхней части бока (у многих карповых- до переднего края основания спинного плавника), а цифра внизу – чешуи под боковой линией до самой нижней точки чешуйного покрова на боку (у многих до основания переднего луча брюшного плавника). Чтобы получить максимальное число продольных боковых рядов чешуи, нужно к сумме верхнего и нижнего чисел прибавить единицу, которая соответствует чешуе, лежащей в боковой линии, если боковая линия есть. На рис. 3 в боковой линии (ряд чешуи с продольными трубочками) по левому боку рыбы 44 чешуи, над боковой линией 8, под боковой линией 5. Боковую формулу здесь надо записать так: 1.1. 4-4 8/5.
Следует указывать количество чешуи по боку хвостового стебля, т. е. число чешуи, лежащих между точками f и d.
Американские авторы, помимо формулы количества чешуи над и под боковой линией, дают общее количество чешуи между началом спинного и брюшного плавников.
Большое систематическое значение имеет количество лучей или косточек в плавниках, причем надо определять главным образом число мягких или ветвистых лучей, так как число жестких неветвистых или простых лучей подвержено слабому колебанию у представителей не только одного вида, но даже и целого рода. Принято просчитывать лучи в первую очередь в спинном и анальном плавниках, и лишь иногда и в других плавниках. Если перед исследователем новое, еще не ^ описанное семейство, новый род, новый вид, то нужно просчитать лучи и простые и ветвистые во всех плавниках. В некоторых иностранных ихтиологических работах, особенно в американских, количества лучей в плавниках даются в виде сумм от сложения числа лучей простых и ветвистых.
Число лучей в спинном плавнике. Спинной или дорсальный плавник принято обозначать буквой D (сокращенное dorsalis). Неветвистые, нерасчлененные (продольно) лучи обозначаются римскими, а ветвистые (расчлененные)- арабскими цифрами. Простые лучи бывают жесткие (колючие и костяные), как у судаков, и мягкие, как у налима. У колюшек перед спинным плавником бывают колючки (иглы), их считают отдельно от числа лучей в плавнике.
Пример: D III 9 .означает, что в спинном плавнике три луча Простых и девять ветвистых; запятой римское и арабское числа здесь ре отделяются. Необходимо помнить, что нередко первый Простой луч почти незаметен и может быть отделен только иглой или скальпелем, а последний ветвистый луч иногда имеет настолько глубокое расщепление, что по внешнему виду его можно принять за два. Между тем подобные ветвистые лучи как в спинном, так и в анальном плавниках расположены на одном общем основании, на одной общей косточке, и поэтому такие лучи принято считать за один, хотя в ведомости лучше обозначать, как 11/2 (полтора); так мы будем накапливать материал по изменчивости числа лучей. Установлено закономерное колебание числа плавниковых лучей в зависимости от географического распространения рыб. А. И. Букирев (1938) считает, что у пеляди (Coregonus peled) при определении количества лучей в плавниках нужно принимать во внимание и крайний (задний) луч, который по наружному виду хотя и представляется недоразвитым, но имеет свое особое основание.
В определении первого простого луча спинного плавника есть разногласия: одни ихтиологи первым лучом считают луч, вышедший из-под кожи; другие, отыскивая первый луч, углубляются под кожу. Полагаю, что более правильным нужно прижать первый способ. В описании плавников нужно пояснить, каким способом определяли число лучей.
Когда рыба имеет два спинных плавника, то лучи первого плавника обозначаются римскими цифрами, если они жесткие (у судака), а лучи второго плавника: простые- римскими, ветвистые-арабскими. Если спинные плавники резко разобщены, то между цифрами, относящимися к плавникам, ставится запятая. Например: D XIII, III 13 значит, что в первом спинном плавнике простых лучей 13, во втором-3 простых и 13 ветвистых. Для налима, у которого оба спинных плавника с мягкими лучами, число лучей обозначается так: I D II – 15, II D 68-82. У наваги (Eleginus) 3 спинных плавника. Формула лучей в D: I D 13, II D 18-20, III D 18-19.
При просчете лучей в анальном или подхвостовом плавнике, который обозначается буквой А (сокращенное analis), надо пользоваться теми же указаниями, что при счете лучей спинного плавника.
В таблице измерений следует для некоторых рыб, как я уже говорил, давать число лучей и других плавников: грудных-Р (pectoralis), брюшных V (ventralis) и хвостового С (caudalis). Точный просчет лучей этих плавников затруднителен, особенно грудных и брюшных плавников, так как нижние мягкие лучи грудных и верхние брюшных плавников обычно мелки и тесно расположены. Для хвостового плавника обычно указывают число лучей хвостовой выемки или развилки, т. е. два длинных простых луча верхней и нижней лопастей плавника плюс все ветвистые лучи. Помимо этих лучей есть простые лучи на верхнем и нижнем боках хвостового плавника, но эти лучи просчитывать крайне трудно. Лучей в С у сигов VI+17+VI, что значит: в хвостовом плавнике по 6 простых лучей в обеих лопастях (6 в верхней и 6 в нижней) и 17 ветвистых лучей в обеих лопастях, но обычно принимают во внимание лишь 19 лучей хвостовой развилки.
Рис. 5. Глоточные зубы карповых (по Юдкину, 1951):
1-однорядные, формула 6-5 (плотва); 2-двухрядные, формула 3.5-5.3 (жерех); 3-трехрядные, формула 1.1.3-3.1.1 (сазан).
Формула глоточных зубов карповых рыб (рис. 5). Эти зубы расположены на пятой (последней) жаберной дуге. Нижнеглоточную кость с зубами извлекают через наружное жаберное отверстие. Для этого нужно ножом отделить верхний край ключицы (cleithrum), прилегающей (снаружи) к заднему краю жаберного отверстия, и вынуть эту кость с глоточными зубами. Глоточные зубы у карповых бывают однорядные, двухрядные и трехрядные; для тех и других применяются |определенные формулы. Число зубов одной стороны отделяется от числа зубов другой горизонтальной линией; если зубов не один ряд, то ближе к черте дают числа зубов, расположенных на внешнем (нижнем) крае кости, а затем числа других рядов. Для однорядных, например, 6-5, т. е. с левой стороны 6 зубов, с правой-5 (плотва); для двухрядных, например, 3.5- 5.3, т. е. на левой стороне в одном ряду три, в другом 5 зубов, с правой- в одном ряду 5, в другом-3 (жерех); пример формулы трехрядных зубов: 1. 1.3-3. 1. 1 (сазан). Если наблюдается рыба в момент смены зубов, когда наряду с новыми имеются и старые зубы, то последние считать не следует.
Число жаберных тычинок (spinae branchiales сокращенно- sp. br.) просчитывается обычно на первой дужке (об этом признаке более подробно см. в пояснениях схемы измерения сигов). Н. В. Европейцева (1939) исследовала строение жаберного аппарата (строение жаберных тычинок) у окуня, .судака и ерша разного возраста и сделала такой вывод: «можно сказать, что тут мы имеем стройный переход от хищного типа питания взрослого судака к полухищному у окуня и до мирного типа питания у ерша с соответствующей каждому типу питания структурой жаберного аппарата». У молодых судаков (длина до 5-6 см), которые питаются преимущественно мелкими ракообразными и лишь переходят на питание рыбой, жаберные тычинки сначала длинные и тонкие постепенно переходят в более утолщенные; при длине судака 30-38 см, когда основной пищей его служит рыба, жаберные тычинки имеют вид толстых зубчатых бугорков. Подмечена также связь структуры жаберного аппарата с характером пищи у окуня и ерша. Автор делает общий вывод, что «у рыб, имеющих жаберные тычинки в раннем возрасте при наличии планктонного питания, по-видимому, нужно всегда ожидать превращения их в бугорки или частичной редукции с переходом на вполне хищный образ Жизни и превращения функции отцеживания мелких организмов из воды в функцию удержания крупных объектов питания и проталкивания их к пищеводу».
Е. С. Кожина (1958), изучавшая структуру жаберных аппаратов рыб, установила многие детали процесса изменения жаберных тычинок хищных рыб. Особый интерес представляет Постепенное превращение жаберных тычинок щуки из палочкообразных тычинок (у мальков) в шиповатые, грубые утолщения (у взрослых); такие утолщения хорошо помогают щуке удерживать схваченную ею жертву.
Иногда нужно просчитывать число жаберных лучей (radii branchiostegi), поддерживающих жаберную крышку. Хотя этот признак довольно постоянен, но его приводят многие авторы.
Число позвонков (vertebrae) считают без уростиля, принимая его за часть последнего позвонка, или с уростилем- как последний позвонок, который у разных рыб имеет различную форму. Поэтому всегда надо упоминать, как производился счет позвонков. Л. С. Берг (1948) писал, что последний зачаточный позвонок со всеми гипуралиями нужно принимать за один позвонок.
Число позвонков имеет важное значение при работах по расовому изучению рыб. Гейнке (Heincke, 1898) положил в основу расовой группировки сельдей число позвонков. Многие русские и иностранные авторы при изучении рыб, особенно при расовом исследовании, пользуются просчетом позвонков. Однако число позвонков значительно варьирует в зависимости от температуры и солености воды в период образования позвонков.
Такие признаки, как число чешуи в 1.1., лучей в плавниках. жаберных тычинок, жаберных лучей, число позвонков и все другие признаки, определяемые путем просчета, называются признаками меристическими, или счетными, а признаки, определяемые при помощи промеров, называются пластическими или измерительными. В систематическом отношении при расовом изучении рыб большое значение имеют пластические признаки, но расовое значение придается и счетным признакам, на которые указывает целый ряд ихтиологов. Например, В. В. Петров (1926) признает, что наиболее надежными признаками при разграничении мелких таксономических единиц уклеек являются признаки меристические.
Осью тела мы называем (несколько нарушая геометрическую точность) ту прямую линию, которая начинается от вершины рта, вернее-от передней точки ротовой щели, при закрытом рте, и заканчивается у корней средних лучей хвостового плавника. Эту линию в таблице промеров легко изобразить графически, обозначив ее направление в зависимости от глаза, условно изображенного в таблице промеров в виде кружка: ось тела пересекает диаметр глаза-рот конечный, ось тела проходит под глазом-рот нижний, ось тела проходит над глазом-рот верхний.
Длина рыла (an), или предглазничный отдел, предглазничное пространство головы, предглазье-от вершины рыла до переднего края глаза, до переднего наружного края глазного яблока. У молодых рыб рыло длиннее, чем у взрослых. Кроме того, есть формы (морфы) длиннорылые и короткорылые.
Длина рыла сильно изменяется у лососей в нерестовый период, а потому для подробных промеров нужно брать лососей с нормально развитыми челюстными костями.
Диаметр глаза (np),если не оговорено особо, берется горизонтальный (продольный). Измеряется собственно диаметр роговицы; веки, если они есть, в расчет не принимаются. Горизонтальный диаметр глаза нередко называется и длиной, глаза. Иногда берется и вертикальный диаметр глаза.
Заглазничный отдел головы (ро), заглазничное пространство- от заднего края глаза до наиболее удаленной от конца рыла точки жаберной крышки; жаберная перепонка 🙁membrana branchiostega), окаймляющая сзади жаберную крышку, в расчет не принимается (у щук, например, эта перепонка очень широкая); не принимаются в расчет и шипы, которые бывают на жаберных крышках (например, у окуня и ерша).
Длина головы (линия ао)- расстояние сбоку от вершины рыла (при закрытом рте) до заднего, наиболее удаленного края жаберной крышки (без жаберной перепонки).
Для рыб, у которых дается толщина тела, нужно указать и Толщину головы; определение толщины головы у таких рыб, как бычки, обязательно.
В работах чехословацких ихтиологов указывается для карповых толщина тела, толщина головы и толщина хвостового стебля.
У многих рыб величина, определяющая длину головы, не является арифметической суммой длины рыла, диаметра глаза и, заглазничного пространства, так как эта сумма графически представляет ломаную линию, линия ао-прямая между крайними точками этой ломаной линии и потому меньше нее. При заполнении таблицы промеров приведенным замечанием можно пользоваться для контролирования цифр промеров указанных частей головы.
Высота головы у затылка (lт). Верхняя точка берется там, где оканчивается череп, нижняя, противоположная ей-по вертикали (линия 1т). У многих рыб верхняя точка находится там, где оканчивается чешуйный покров, но у некоторых рыб, например у семейства кефалевых (Mugilidae), чешуя заходит и на лоб; у таких рыб нужно определить задний конец верхнезатылочной (supraoccipitale) кости. У осетровых линия высоты головы проходит впереди первой спинной жучки. Иногда высота головы берется и через середину глаза, что оговаривается особо.
Ширина лба или межглазничное пространство, межглазничный промежуток-расстояние между глазами сверху, т. е. ширина черепа между глазами (см. рис. 3, справа).
Наибольший обхват тела измеряется сантиметровой лентой в месте наибольшей толщины и наибольшей высоты тела, не беря в расчет плавников. Эту величину нужно записывать в таблицу промеров в сантиметрах, так как более точное определение здесь почти недостижимо.
Наибольшая толщина тела – наибольшее расстояние между боками, Это необходимо для миног, осетровых, угрей, сомов, налимов, не лишние приводить эту величину и для других рыб. Но следует помнить, что обхват и толщина тела сильно изменяются.
Наибольшая высота тела (gh)-расстояние от самой высокой точки спины до брюшка по вертикали; плавники, а у осетровых и костяные щитки в расчет не входят.
Высота тела – величина довольно изменчивая: у старых особей (семейства карповых) тело (относительно к длине) выше, чем у молодых; кроме того, среди рыб нередки морфы: высокотелая (morpha elata) и низкотелая, удлиненная (m. elongata). Когда имеется материал плохо законсервированный (вздутое или слишком ослабнувшее брюшко), точно установить наибольшую высоту тела трудно.
Наименьшая высота тела (ik) обычно находится близ основания хвостового плавника, часто на середине хвостового стебля. Многие авторы эту величину называют высотой хвостового стебля.
Антедорсальное расстояние, антедорсальное пространство (aq) – от вершины рыла до основания первого луча спинного плавника. Эта линия является прямой, соединяющей обе указанные точки. Для характеристики кривой, идущей от рыла до спинного плавника через лоб и спинку рыб, следует это расстояние измерять по спине лентой или ниткой. Но обычно такое измерение игнорируется, и профиль спины дается описательно: спина горбатая, линия спины правильно дугообразная и т. п.
Постдорсальное расстояние, постдорсальное пространство (rd) -от вертикали заднего конца основания спинного плавника до основания хвостового плавника, считая по середине тела. Если плавников два или три, эту линию откладывают от конца основания первого плавника, если он отделен от других; если плавники сращены, постдорсальное расстояние берется от вертикали второго плавника, но это в таблице должно быть оговорено.
Расстояние от конца рыла до брюшного и анального плавников (антевентральное и антеанальное) и расстояние от брюшного и анального до основания хвостового (поствентральное и постанальное расстояния) для карповых рыб брать не обязательно.
Длина хвостового стебля (fd)-расстояние от вертикали заднего края основания анального плавника до основания хвостового плавника или до конца чешуйного покрова. Откладывают эту линию по середине тела рыбы. У осетровых длина хвостового стебля определяется иначе (см. ниже).
Длина основания D – длина спинного плавника (qs) – от основания переднего, хотя бы зачаточного, луча до основания последнего луча или до конца перепонки (если она есть) спинного плавника. Если спинных плавников не один – показать длину и другого (или других); при слитых спинных плавниках (у ерша) определить расстояние до конца последнего луча каждого плавника.
Наибольшая высотa D (tu) – высота спинного плавника -высота наибольшего луча этого плавника; если луч согнут, то его нужно расправить. У хариусов (сем. Thymallidae) взрослые самцы имеют более высокий дорсальный плавник. Изменение D у хариусов описано В. К. Черновым (1934).
Длина основания А, длина анального плавника,- условия те же, что и при намерении длины основания D.
Наибольшая высота А, высота анального плавника,- условия те же, что при измерении высоты D.
Длина Р (vx) -и длина V (zz1), т. е. длина грудных и длина брюшных плавников,- от передней линии их прикрепления до вершины наиболее длинного луча. У наваги второй луч брюшного плавника вытянут в нить; длину этой нити следует показывать отдельно, не включая ее в общую величину длины V.
Длина верхней лопасти хвостового плавника (С)- длина наибольшего луча верхней лопасти хвостового плавника.
Длина нижней лопасти С-длина наибольшего луча нижней лопасти хвостового плавника.
Для определения величины хвостовой выемки или хвостовой округленности можно измерить средний луч хвостового плавника. Этот способ недостаточно удовлетворителен в тех случаях, когда степень хвостовой выемки приводится как признак систематической группировки, например у ельцов, гольянов, пескарей; более точным нужно считать отношение линии от наружного края средних лучей С до линии, соединяющей концы верхней и нижней лопастей плавника, к этой последней линии.
Расстояние между Р и V–расстояние между основаниями грудного и брюшного плавников, передняя часть брюха, промежуток между грудными и брюшными плавниками – определяется от передней точки прикрепления одного плавника до передней точки прикрепления другого плавника (линия vг). Есть рыбы, у которых грудные плавники лежат над брюшными или позади брюшных (тресковые), тогда расстояние между Pи V определять не нужно.
Расстояние между V и А, – расстояние между брюшным и анальным плавниками, этот промежуток называют также задней частью брюха.
У гольянов нужно брать, кроме того, толщину хвостового стебля, а у рыб, имеющих усики, следует измерить их и сравнить, например, с диаметром глаза. При описании подуста дается измерение расстояния между углами рта, причем это расстояние выражается в процентах длины головы.
Образец таблицы промеров карповых см. в приложении I. На первой и четвертой страницах таблицы дают замечания к тому или иному номеру: аномалии, уродства, прижизненная окраска глаз, окраска и форма плавников и т. д. Для карасей, ельцов и гольянов обязательно указывают число тычинок на первой жаберной дуге, а для пескарей – длину и число усиков и расстояние от конца рыла до анального отверстия, потому что положение анального отверстия у пескарей сильно варьирует.
Усики следует измерять при изучении всех представителей усатых рыб (храмуль, усачей, маринок, османов, усатых гольцов, вьюнов и др.).
Если невозможно определить с нужной точностью какую-либо величину, в таблице в соответствующей графе ставится знак вопроса, например 17 (?) или са (causa-около). Делать это можно лишь в том случае, когда неточность не нарушает общего характера вычисления, т. е. когда она незначительна.
Образец ведомости индексов для карповых рыб см. в приложении II.
Заметим, что многие иностранные авторы, прежние и современные, держатся обычая выражать отношения между частями тела рыб в кратных показателях, а не в процентах.
По схеме карповых можно измерять и чукучана (род Саtostomus сем. Catostomidae), но для него обязательно всегда указывать количество тычинок на первой жаберной дужке. Подсчет количества жаберных тычинок на первой дужке нужно вести у всех рыб, где эти тычинки выражены. Помимо количества тычинок, надо указывать длину наибольшей тычинки и всей жаберной дужки, а в ведомости индексов выражать длину наибольшей тычинки в % длины дужки.
К семейству карповых в водах Союза ССР относится более 50 родов с более чем 200 видами и разновидностями рыб, между которыми немало обладающих чрезвычайно большой изменчивостью; поэтому в описании таких рыб необходимо пользоваться, помимо общей схемы измерений, и частными указаниями, имеющимися в литературе. Например, при изучении пескарей (род Gobio), среди которых известно много систематических форм, следует дополнить схему карповых такими признаками: количество темных пятен на теле, длина усика, расстояние от усика до заднего края praeoperculum, толщина хвостового стебля, расстояние от А до ануса, расстояние от V до ануса, длина наибольшей жаберной тычинки, длина первой жаберной дужки. Толщина хвостового стебля у пескарей измеряется у заднего края анального плавника. Расстояние от V до ануса измеряется от заднего края основания V.
Помимо того, следует в примечаниях или в ведомости промеров условными знаками (+ или -) указывать следующие признаки: горло голое или покрытое чешуей, угол рта достигает или не достигает вертикали переднего края глаза, последний простой луч D утолщен или не утолщен, профиль рыла с выемкой или сферический, пятна на D, пятно темное за жаберным отверстием, когда темных пятен на боках нет, то обычно есть темная полоса.
Многие из признаков пескарей подвержены сильным колебаниям, вследствие чего систематика этой группы рыб представляет большие трудности, и полного описания всех пескарей даже для водоемов СССР еще не сделано; между тем представители рода Gobio распространены и в Китае, и в Японии.
На основании изменчивости признаков Л. С. Берг установил часто встречающиеся типы вариации пескарей (morpha): m. brevicirris и m. longicirris(коротко- и длинноусые пескари); m, prosopyga и m. katapyga (у первых анус отодвинут к основанию брюшного плавника, у вторых – к основанию анального плавника), m. obtusirostris и m. longirostris(короткоры-лые и длиннорылые).
Измерение рыб лососевых
(Salmonidae)
Систематика рыб семейства лососевых вследствие большой изменчивости этой группы представляет собой одну из наиболее трудных и наиболее невыясненных задач современной ихтиологии. До сих пор не вполне установлены границы даже родов »того семейства, не говоря о границах видов и других, более мелких таксономических единиц. Род Thymallusвыделен в особое семейство Thymallidae. Только начата разработка систематики рода Salvelinus.
При промерах лососей обычно пользуются схемой Смитта, Который дал богатейший материал по систематике лососевых. Но схема измерений лососевых, предложенная Смиттом, оставаясь в общем необходимой, требует изменений и пояснений. Мы приводим здесь схемы измерений лососей и сигов, составленные в основном по схемам Смитта, но мы их сократили по Сравнению со схемами,, которые изложены в 3-м издании нашего Руководства (1939). Целесообразность такого/сокращения изложена мной ранее (Правдин, 1951).
Измерение лососей
За основную длину тела лососей принимается расстояние от вершины рыла, т. е. от передней, наиболее удаленной точки тела при закрытом рте до наружного конца средних лучей хвостового плавника (на рис. 6 ab). К этой длине по Смитту относят промеры других частей тела. Передней точкой у одних лососей является передний край верхней челюсти, у других – край нижней челюсти. Следует приводить и длину без хвостового плавника (без С). Оставляя за термином «длина тела» то значение, какое придал ему Смитт, можно вместе с тем предложить ввести и для лососей длину туловища.
Длина всего тела или предельный размер у лососей исчисляется от конца рыла до середины линии, соединяющей концы верхней и нижней лопастей хвостового плавника (аb).
Число чешуи в боковой линии у лососей просчитывать весьма трудно, поэтому, когда чешуя очень мелкая, можно ограничиваться определением числа чешуи в 1/10 длины тела, отложив эту величину по боку тела впереди спинного плавника, повыше боковой линии. Можно также определить число чешуи от заднего края жирового плавника по боковую линию, включая сюда и чешуйку боковой линии. Но оба эти определения не дают надежных результатов.
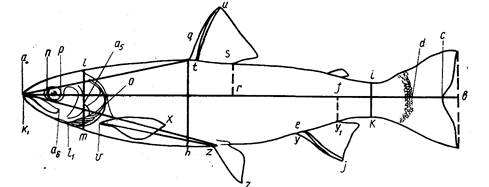
Рис. 6. Схема измерений рыб лососевых, по Смитту, с изменениями:
аb-длина всей рыбы; ас-длина по Смитту; ad-длина без С; od-длина туловища; an-длина рыла; пр-диаметр глаза (горизонтальный); аа5– длина средней части головы; ао-длина головы; ро-заглазничный отдел головы; lm-высота головы у затылка; ширина лба (как у карповых); ad6 – длина верхнечелюстной кости; k1l1 – длина нижней челюсти; qh – наибольшая высота тела; ik – наименьшая высота тела; aq – антедорсальное расстояние; rd – постдорсальное расстояние; аz- антевентральное расстояние; ay – антеанальное расстояние; fd – длина хвостового стебля; qs – длина основания В; fu – наибольшая высота D; уу1 – длина основания A; ej-наибольшая высота А; ох-длина Р; zz1-длина V; vz-расстояние между Р и V; гу – расстояние между V и Л.
Достаточно измерить горизонтальный диаметр глаза, причем веки в расчет не принимаются.
У лососей нужно указывать также число и характер жаберных тычинок (тычинки заостренные, тупые, булавовидные), число жаберных лучей, пилорических придатков (последнее- весьма трудная задача), описать глубину хвостовой выемки.
При промерах рыб лососевых нужно пользоваться таблицей, данной в приложении III. Ведомость индексов см. в приложении IV.
По схеме промеров лососей измеряют гольцов (Salvelinus), род Hucho, род Brachymystax.
Измерение сигов
Род сигов (Coregonus), так же как и лососи, широко распространен и обладает свойствами давать множество местных географических и экологических форм. Трудно поддается систематической группировке. Только подробные промеры и многочисленные индексы, проанализированные методом вариационной статистики с учетом биологических особенностей, могут привести к надежным результатам по разграничению разных, даже самых мелких групп (рас и форм).
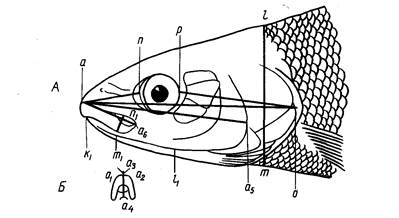
Рис. 7. Схема измерений головы сига, по Смитту, с изменениями:
А- голова сига: ао – длина головы; aa5 – длина средней части головы; aп – длина рыла; ро – заглазничный отдел головы; aa6 – длина верхнечелюстной кости; m1n1– ширина верхнечелюстной кости; k1l1 – длина нижней челюсти; пр – диаметр глаза горизонтальный; lm – высота головы у затылка. Б-рыльная площадка: а1а2–ширина площадки рыла; а3a4– высота площадки рыла.
Смитт для промеров сигов предложил особую схему, которая очень близка к схеме промеров лососей. Особенность схемы измерений сигов – за длину тела сигов при измерениях берут расстояние от переднего наружного края верхнечелюстной кости (maxillare), от точки а (рис. 7), до наружного края средних лучей хвостового плавника. Напомним, что передний край верхнечелюстной кости хорошо обозначается, если оттянуть вперед и вбок верхнечелюстную кость за ее задний край. У молодых сигов передний край maxillare при плотно закрытом рте почти совпадает с передней точкой тела (разница 0,5-1 мм).
При измерении нижней челюсти сигов за задний конец ее принимают место, где нижняя челюсть сочленяется с квадратной костью, место это заметно, если нижнюю челюсть отодвинуть сильно вниз. Для сигов, кроме того, нужно давать измерения площадки рыла, т. е, передней наружной части верхней челюсти, расположенной между верхнечелюстными костями (указать высоту и ширину площадки). Приводят и меристические признаки: число чешуи в боковой линии (как у карповых), жаберных тычинок, жаберных лучей и число позвонков.
Значение жаберного аппарата в систематике сигов выяснено многочисленными работами немецкого ученого А. А. Тинемана (1928) и финского ихтиолога Ярви (1928). Промеры головы сигов показаны на рис. 7.
Рис. 8. Первая жаберная дуга сига (по Правдину, 1939):
ab – длина нижней части дуги, bс – длина верхней части дуги.
Не у всех сигов направление рыльной площадки одинаковое: у одних площадка рыла выпуклая, делающая рыло овальным и острым; у других – вертикальная, делающая рыло вертикально-усеченным; у третьих площадка скошена вниз и назад. Направление площадки, по-видимому, характерно для отдельных групп сигов.
Форма жаберных тычинок у разных групп сигов тоже различна: гладкие, пильчатые, с зубчиками (на рис. 8 представлена первая жаберная дужка сига), с добавочными веточками. Есть сиги с круглыми, .цилиндрическими тычинками и с тычинками плоскими. У некоторых сигов против длинных тычинок, на другой стороне жаберной дужки, бывает ряд довольно развитых добавочных тычинок, зубчиков.
Неодинакова и форма плавников: есть плавники с острыми и тупыми концами, есть с глубокой и со слабой выемкой.
Окраска сигов весьма разнообразна. Мы не придаем ей систематического значения, но надо обращать внимание на пятнистость, которая наблюдается на голове и плавниках некоторых сигов.
Зрачки у сигов бывают угловатые (спереди) и более круглые. | Все упомянутые особенности нельзя считать надежными систематическими признаками. Это – лишь вопросы, которые пока не получили надлежащего освещения и над которыми следует доработать.
По схеме сигов измеряют рыб рода Stenodus(белорыбицы, нельмы) и семейства Thymallidae (хариусовые).
По описанию признаков хариусов имеются две большие работы А. Н. Световидова (1931, 1936). В этих работах можно найти Многочисленные указания и пояснения, относящиеся к методам изучения систематики и биологии хариусов. Высоту спинного Плавника нужно измерять в двух местах: «в передней части длинного плавника по последнему неветвистому лучу и в задней части плавника по четвертому ветвистому лучу от конца плавника». При подсчете простых лучей в D и в А А. Н. Световидов считал и те лучи, которые скрыты в коже рыбы.
Ряпушки (Coregonus albula, Coregonus sardinella), которые относятся к роду сигов, измеряются, конечно, по схеме сигов.
В приложениях III и IV даны образец таблицы для записи промеров и образец ведомости для записи индексов для лососей и сигов (вместе). Первая таблица составлена с таким расчетом, чтобы ее можно было использовать для записи промеров лососей и сигов и других рыб, измеряемых по таким же схемам. В таблице есть графы, которые не всегда нужно заполнять (например, лучей в Р. V и С, чешуи в 1/10 длины тела), и, кроме того, введена графа для записи длины расстояния от конца рыла до переднего края maxillare, т. е. до точки а. Эта величина в дальнейшем может служить той поправкой, которая дает возможность перевести наши промеры длины тела, рыла, головы, антедорсального, антевентрального и антеанального расстояний на численные данные таких промеров, которые велись бы при игнорировании величины расстояния от конца рыла до maxillare.
Ведомость для записи индексов можно составить так, чтобы она включала и все меристические признаки и таким образом | выла бы пригодна и для дальнейшей обработки.
Чтобы сравнить описываемую форму с ранее установленной, нужно пользоваться и другими промерами, которыми пользовались авторы, описавшие ее.
Измерение рыб корюшковых
(Osmeridae)
Корюшек, как и всех представителей семейства Osmeridae, обычно измеряют по схеме промеров лососей, т. е. по схеме Смитта. Более поздние работы по систематике корюшек (Тюрин, 1924, Петров, 1925) показали необходимость приводить для корюшки и другие признаки. Для изучения форм корюшек следует придерживаться определения таких признаков (этот порядок следует сохранить для таблицы промеров): 1) пол и состояние половых продуктов, 2) вес, 3) возраст, 4) длина всей рыбы, 5) длина тела по Смитту (от переднего края нижней челюсти при закрытом рте до конца средних лучей С), 6) длина тела без С, 7) длина туловища. Признаки счетные: 8) число чешуи, прободенных боковой линией, 9) число поперечных рядов чешуи, 10) лучей в D, 11) лучей в А, 12) число тычинок на первой жаберной дужке. Из перечисленных меристических признаков наиболее существенные: число чешуи в 1.1., число лучей ветвистых и неветвистых в D, число ветвистых лучей в Л и число жаберных тычинок. У корюшек бывает значительная разница в количестве простых (неветвистых) лучей спинного плавника (Петров, 1925).
Признаки пластические: 13) длина рыла, 14) диаметр глаза, 15) заглазничное пространство, 16) длина головы, 17) высота головы у затылка, 18) ширина лба, 19) длина верхней челюсти, 20) длина нижней челюсти, 21) наибольший обхват тела, 22) наибольшая толщина тела, 23) наибольшая высота тела, 24) наименьшая высота тела, 25) антедорсальное расстояние, 26) постдорсальное расстояние, 27) длина хвостового стебля, 28) расстояние от конца D до жирового плавника, 29) длина D, 30) высота D, 31) длина Р, 32) длина V, 33) длина Л, 34) высота А, 35) расстояние между Р и V, 36) расстояние между V и А.
Некоторые из перечисленных пластических признаков, по замечанию В. В. Петрова, систематического значения не имеют. К ним автор относит: наименьшую высоту тела, длину D, длину Р, длину А, высоту А, длину челюстей. Наиболее существенными признаками он считает: величину антедорсального и постдорсального расстояний, расстояние V-А, длину V и ширину лба.
При описании корюшек нужно давать характеристику их зубов. В.В. Петров подробно рассмотрел и биометрически проанализировал многочисленные признаки корюшек: невской, беломорской (сорокской), дальневосточной, онежской и енисейской.
Подобную работу он проделал в отношении систематики ладожской корюшки (Петров, 1926).
Измерение рыб осетровых
(Acipenseridae)
Схема промеров осетров изображена на рис. 9 и приводится в приложении V.
Длина всей рыбы, или абсолютная длина тела, измеряется от вершины рыла до вертикали конца верхней лопасти хвостового плавника (линия аb). С этой величиной сравнивают все другие промеры (у осетровых рыб верхняя лопасть хвостового плавника представляет продолжение тела, продолжение хвостового стебля рыбы). Но вместе с тем рекомендуется указывать и длину тела от конца рыла до конца средних лучей наиболее глубокой средней части вырезки хвостового плавника. Мы указываем, кроме того, длину тела до последней жучки или до корней средних лучей С (линия ad).
Нужно давать описание формы усиков (усики в разрезе круглые, сплющенные, с листовыми придатками, как, например, у белуги- Huso huso и без придатков, как у калуги-Huso dauricus); описание жаберных перепонок-приращены ли они к межжаберному промежутку (isthmus), как у рыб рода Acipenser (осетры), или срастаются между собой, образуя свободную складку, как у рыб рода Huso (белуги).
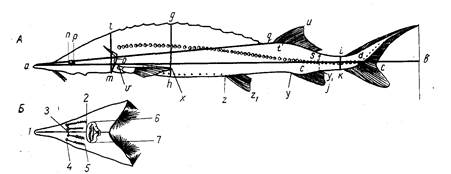
Рис. 9. Схема измерений осетровых рыб (Acipenseridae):
А – аb – длина всей рыбы; ас – длина до конца средних лучей С; ad – длина до корней средних лучей С; od-длина туловища; an-длина рыла; nр-диаметр глаза (горизонтальный); ро-заглазничный отдел головы, ao – длина головы; lm – высота головы у затылка; gh – наибольшая высота тела; ik – наименьшая высота тела; fd – длина хвостового стебля; aq – aнтедорсальное расстояние; аz-антевентральное расстояние; ау-антеанальное расстояние; qs – длина основания D; tu – наибольшая высота D; уу1–длина основания A; ej-наибольшая высота A; vx-длина Р; zz1– длина V; vz-расстояние между Р и V; zу – расстояние между V и А. Б. Голова снизу: /-2-расстояние от конца рыла до хрящевого свода рта; 1-3 – расстояние от конца рыла до средних усиков; 4-5 – длина наибольшего усика; б-7-ширина рта.
Морфологическое описание осетровых лучше делать по взрослым, вполне сформировавшимся особям, так как некоторые признаки у молодых осетровых довольно непостоянны.
Напомним, что жировая подушка плавников осетровых при измерениях в расчет не принимается.
В 1934 г. вышла ценная работа Н. И. Сальникова и С, М. Малятского (1934) по биометрическому изучению белуги Азово-Черноморского бассейна.
Исследованиями Азербайджанской научной рыбохозяйственной станции еще более 10 лет тому назад установлено, что в Каспийском море имеется по меньшей мере три морфологически и географически обособленные формы осетра – волжский осетр, сефидрудский и куринский (Голованов, 1940). Известно также, что в Азовском море обитают два стада севрюги – донское и кубанское, имеющие различные места нереста и различный темп роста (Чугунова,1940).
Интересные наблюдения А. Н. Пробатова (1935) над ростом амурской калуги (Huso dauricus) показывают, что этому виду свойственно образование местных форм (жилая и проходная). Кроме того, у калуги, как и у многих других осетровых, есть яровая и озимая формы, на что указывают многие авторы (Берг, Пробатов, Недошивин, Телегин, Лукин, Трусов и др.). При изучении систематики осетровых нужно помнить, что среди них часто встречаются помеси.
Ведомость индексов можно составлять по образцу приложения II с дополнением промеров, типичных для осетровых.
Измерение рыб сельдевых
(Clupeidae)
У сельдей нет боковой линии, поэтому у них нужно просчитывать число поперечных рядов чешуи. Кроме того, у многих сельдей чешуйный покров заканчивается двумя очень длинными чешуйками, далеко заходящими за корни средних лучей хвостового плавника. Эти чешуйки (alae) не принимаются в расчет ни при счете поперечных рядов чешуи, ни при измерении длины тела от переднего края рыла до конца чешуйного покрова.
При группировке сельдей большое систематическое значение придают числу и характеру жаберных тычинок. При разграничении отдельных видов и подвидов сельдей этот признак весьма существен, но для характеристики рас сельдей его недостаточно, так как количество тычинок с возрастом сельдей изменяется.
Видовое определение сельдей по количеству жаберных тычинок особенно трудно у молоди. Б. И. Диксон (1905) указывает, что у мальков сельди черноспинки, как правило, меньше жаберных тычинок, чем у взрослых особей того же вида: при длине мальков 18 мм тычинок на первой жаберной дужке было 10- 11, при длине мальков 20 мм-11-13 тычинок, при длине 22,5 мм – 20 тычинок; у взрослых особей черноспинки жаберных тычинок обычно больше 60.
Д. Ф. Замахаев (1944), рассматривая систематическое положение проходных каспийских сельдей, пришел к заключению, что структура жаберных дужек и тычинок, стадии зрелости половых продуктов, темп роста, структура чешуи у среднетычинковых сельдей не имеют систематического значения, и что черноспинка, малотычинковая волжская сельдь и сельдь Берга- « являют собой отдельные возрастные группировки одного вида». Этот вопрос требует дополнительных исследований, так как сущность не в том, считать ли названных сельдей отдельными видами, подвигами или более мелкими таксономическими единицами, а существенны их морфологические и биологические признаки, которые все же имеются.
Локальные формы сельдей (а их множество) в большинстве Случаев имеют хорошо заметные биологические различия (различные сроки нереста, температуры при размножении и темп роста). Обычно можно подметить и морфологические различия. Такие морфологические различия удалось установить А. И. Фролову (1950) у трех стад сахалинской сельди (декастринского, западно-сахалинского и анивского).
Д. К. Третьяков (1938) описал методику изучения сейсмосензорных каналов сельдей. Сельдей консервировали крепким спиртом, высушивали на воздухе в продолжение 2-3 дней, после чего в один из наиболее заметных после просушки сейсмосензорных каналов головы делали инъекцию жидкой цветной или черной тушью. После инъекции голову окончательно высушивали.
В статье А. И. Дехтяревой (1940) имеется определительная таблица для сеголетков сельдей и даны хорошие рисунки семи разновидностей каспийских сельдей.
Описания личиночных стадий рыб все чаще и чаще встречаются в литературе. Здесь снова приходится сослаться на статьи С. Г. Крыжановского, А. И. Смирнова, С. Г. Соина (1951), А, Г. Егорова (1961), К. С. Поповой (1961), М. П. Рыженко (1939), Н. В. Европейцевой (1946, 1949 а и б), Т. С. Расса (1939) И добавочно указать на статью Т. А. Перцевой-Остроумовой (1951). В последней статье даже указана схема промеров личинок. В конце статьи дается определительная таблица для икринок и для личинок сельдей Северного Каспия.
Основными различительными признаками сельдей продолжают считаться жаберные тычинки и позвонки.
У одних сельдей тычинки тонкие и длинные (например, у Caspialosa caspia), у других-грубые, у третьих-с зубчиками М т. п. На 1-й или на 4-й страницах таблицы промеров необходимо записывать характер тычинок, а также отмечать отсутствие или присутствие зубов на челюстях и сошнике (vomer).
Нужно отмечать также форму головы (верхняя линия профиля). Г. П. Булгаков (1926) главным образом на основании формы головы описал новый вид сельди (Caspialosa kisselevitschi). Для Clupea harengus необходимо просчитывать число зубных рядов на сошнике (этот признак, как показывают работы А. И. Рабинерсона, варьирует с возрастом: у старых рыб сошниковых зубов меньше). Просчет позвонков необходим. Этот признак положен Heincke (1898) в основу систематики норвежских сельдей. По количеству позвонков разнятся многие сельди, в том числе по позвонкам отличают мурманскую сельдь (Clupea harengus harengus) от беломорской (Clupea harengus pallasi natio maris-aibi): у первой позвонков 56-59, у второй 50-56 (по Рабинерсону). Число позвонков у сельдей удобно сосчитать даже •в молодом возрасте (в стадии сеголетков при размере от 4 до 6 см., для мурманской сельди). Последним позвонком у сельдей считают тот (Heincke, 1898), который загибается кверху перед началом хвостового плавника и с которым соединены последние рудиментарные позвонки в уростиль (Берг, 1948).
С. В. Аверинцев (1929) предложил вниманию русских ихтиологов новый метод, применяющийся И. Шмидтом и А. Иогансеном при просчете позвонков рыб. С. В. Аверинцев совместно с Н. В. Сыч пользовался таким методом при изучении беломорских и каспийских сельдей, результаты, как говорит С. В. Аверинцев, оказались блестящими. Они приведены ниже в виде простой и понятной таблицы (табл. 1).
Таблица 1
|
Общее число позвонков в теле рыбы |
Число особей (в %) по отдельным видам |
|||||
|
Clupea kessleri |
Clupea volgensis |
Clupea suvorovi |
Clupea saposhnikovi |
Clupea caspla |
Clupea brashulkovi |
|
|
44 |
– |
– |
– |
– |
0,65 |
0,95 |
|
45 |
– |
– |
– |
– |
0,65 |
– |
|
46 |
– |
– |
– |
– |
1,96 |
1,90 |
|
47 |
– |
– |
– |
– |
2,61 |
|
|
48 |
– |
– |
– |
1,75 |
5,23 |
1,90 |
|
49 |
– |
– |
3,70 |
8,77 |
26,79 |
7,62 |
|
50 |
– |
5,88 |
14,81 |
52,63 |
50,32 |
24,76 |
|
51 |
– |
20,59 |
62,96 |
36,24 |
10,45 |
28,57 |
|
52 |
24,44 |
41,17 |
18,52 |
– |
1,90 |
28,57 |
|
53 |
64,44 |
32,35 |
– |
– |
– |
5,71 |
|
54 |
11,11 |
– |
– |
– |
– |
– |
|
Среднее число позвонков у данного вида |
52,87 |
50,95 |
50,24 |
49,53 |
50,78 |
|
Как ни привлекательна простота таблицы, но и здесь необходима вариационная статистика, которая дала бы возможность сказать, насколько существенно различие между средними количествами позвонков сельдей: без такого анализа едва ли следует вести просчет позвонков двух или нескольких близкородственных форм рыб.
Известно, что сельдь оставшаяся и размножающаяся в опресненных заливах, слабо соединяющихся с морем, имеет меньшее количество позвонков.
Нужно указывать присутствие или отсутствие киля на брюшке и число брюшных шипиков, а также число килевых чешуи, если они имеются. При счете килевых чешуи, как отмечает Л. С. Берг (1948), «за чешуи перед брюшными плавниками считаются чешуи, лежащие впереди передней точки основания брюшных; чешуи за этой точкой считаются лежащими за брюшными плавниками».
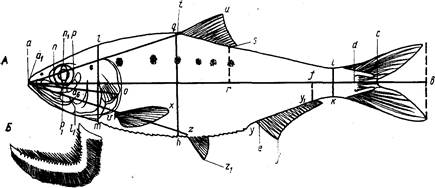
Рис. 10. Схема измерений сельдевых рыб (Clupeidae):
А – аb – длина всей рыбы; ас – длина до конца средних лучей С; ad – длина без С; an – длина рыла; п1р1 – диаметр глаза (вертикальный); ро – заглазничный отдел головы; ао – длина головы; lm – высота головы у затылка; ширина лба (измеряется как у карповых); а1а6 – длина верхнечелюстной кости; аl1-длина нижней челюсти; qh-наибольшая высота тела; ik-наименьшая высота тела; aq-антедорсальное расстояние; rd-постдорсальное расстояние; аz – антевентральное расстояние; ay – антеанальное расстояние; fd – длина хвостового стебля; qs-длина основания D; tu-наибольшая высота D; уу1 – длина основания A; ej – наибольшая высота А; vх – длина Р; zz1–длина V; vz-расстояние между Р и V; zу-расстояние между V и A; dc – длина средних лучей С. Б – жаберная дужка. Верхний ряд – тычинки, нижний – лепестки.
Можно указывать и число пилорических придатков.
Наиболее легкой схемой измерения сельдей является схема, принятая Л. С. Бергом (рис. 10). В ней за длину тела принимается расстояние от вершины нижней челюсти (если она при закрытом рте длиннее верхней) до конца средних лучей хвостового плавника; за всю длину, или абсолютную длину рыбы – расстояние от вершины рыла до перпендикуляра, восстановленного от конца нижней лопасти; диаметр глаза вертикальный, потому что у многих сельдей по горизонтали глаза расположены пеки, закрывающие роговицу глаза (вертикальный диаметр глаза у сельдей обычно равен горизонтальному). Образец таблицы промеров см. в приложении VI.
В ведомостях индексов за основную величину, по которой ведутся вычисления, одни авторы (Суворов, Берг, Киселевич, Булгаков) берут длину тела от вершины рыла до конца средних лучей С (по Смитту); другие (Браунер, Рабинерсон) длину всей рыбы (по Гейнке). В монографии А. Н. Световидова (1952) измерения частей головы выражены в процентах ее длины, а измерения частей тела в процентах всей длины рыбы, кроме сельдей из родов Clupeonella и Alosa, у которых измерения выражены в процентах длины до конца средних лучей хвостового плавника.
Следует заметить, что в систематическом разграничении сельдей Советского Союза нет еще полной согласованности между авторами, поэтому можно считать, что систематика русских сельдей не закончена даже в основных своих формах (в границах вида), не говоря уже о мелких таксономических единицах. Напомним, что в работах по систематике наших сельдей следует исходить из установления тех родов, которых (по Бергу) можно встретить в наших водах.
При описании сельдей приводят окраску (у некоторых имеются характерные пятна), указывают форму рта (у одних верхний, у других полуверхний), присутствие или отсутствие век (у рода Clupea веки зачаточные, а у Caspialosa веки сильно развиты) . Икра у одних (Clupea) прилипающая, у других (у килек) пелагическая. Различают сельдей осенне- и весеннемечущих. Такие расы имеются даже в пределах одного и того же вида (например, у салаки – Clupea harengus membras). Есть расы мелкие и крупные (среди беломорских сельдей). Сельди различаются и по величине диаметра икринок (у килек). Нужно описывать тычинки (толстые, тонкие, длинные, короткие), отмечать, насколько выдается нижняя челюсть за край верхней, а также – как далеко заходит задний край верхнечелюстной кости (по отношению к глазу).
После того как по систематике сельдей проведено несколько новых исследований, можно предложить более полную схему измерений, хотя заметим, что некоторыми авторами по отношению к виду Clupea harengus высказан взгляд, что изучение пластических признаков мало может дать для систематики мелких таксономических групп, на которые разбивается этот вид. Однако без изучения пластических признаков сельдей их систематическая группировка невозможна, это признают и те исследователи, которые не придают пластическим признакам решающего значения. А. И. Рабинерсон (1928) говорит: «анализ пластических признаков, необходим, так как единственный остающийся сверх этого признак-темп роста -сам по себе не может служить в качестве решающего Момента в вопросах систематики».
В небольшой работе А. Браунер (1912) о сельдях Черного и Азовского морей можно найти ценные методические указания как по описательным, так и по измерительным признакам сельдей. Автор, кроме того, использует указания других исследователей (Pallas, 1811; Eichwald, 1838; Normann, 1934; Кесслер, 1874,Antipa, 1905). В статье дается список промеров сельдей. Индексы высчитаны ко всей длине тела и отчасти к длине головы. Браунер дает понятие терминов рыло и морда в отношении сельдей: конец рыла это передний край верхней челюсти, а конец морды – Конец нижней челюсти, которая у сельдей выдается вперед.
К. А. Киселевич в 1923 г. в своей обширной книге о сельдях Привел огромнейшее количество цифр промеров и индексов, как Взрослых, так и молодых (неполовозрелых) сельдей, а также описал опыты учета изменения размеров тела сельдей от консервирования их формалином и спиртом.
А. А. Михайловская составила руководство к определению каспийских сельдей (1941), где можно найти много указаний по Морфологическим признакам сельдей.
Ф. Э. Карантонисом (1938) проведена оригинальная работа по морфологической характеристике мурманских сельдей. Зарисовывали контур голов сельдей. После очистки черепа от кожного покрова и мышц измеряли отдельные части головы. Обработка этих материалов привела исследователя к заключению, что Мурманских (исследованных) сельдей следует разбить на три типа, различающихся по строению костей жаберной крышки: 1) сельди, у которых задняя свободная кромка operculum имеет выемку, нижний свободный край передней кромки operculum не причленяется ни к praeoperculum, ни к interoperculum; 2) сельди, у которых задняя кромка operculum не имеет выемки, а нижний Край передней кромки operculum аналогичен первому типу и 3) сельди, у которых задний край operculum без выемки, а нижний край operculum плотно прилегает к praeoperculum и interoperculum. Каждому из названных типов соответствует характерное строение плечевого пояса и разная форма головы. Череп у первого типа широкий и низкий, верхняя челюсть длинная и более массивная; у второго типа голова небольшая, более стройная и более симметричная; у третьего типа голова массивная и более высокая. Локализация этих трех типов также различна. Сельди первого типа придерживаются преимущественно береговых вод. Сельди второго типа, обладающие большой подвижностью, распространены более широко: «они повсеместно встречаются в Баренцевом море, как в открытой его части, так и в прибрежных водах». Сельди третьего типа обнаружены преимущественно в третьей и во второй струях Нордкапского течения.
Сообщенный Ф. Э. Карантонисом метод различия мурманских Сельдей по строению костей жаберной крышки может быть окажется полезным при изучении систематики не только мурманских, но и других сельдей и даже других семейств рыб.
При изучении сельдей следует знакомиться с размерами » окраской икры (см. работы Т. С. Расса, 1939; Перцевой, 1940 и др).
В упомянутой монографии А. Н. Световидова по сельдевым имеется много ценных указаний и по методике исследований молодых сельдей.
Схемой измерений сельдей можно пользоваться и для измерения рыб семейства Engraulidae, к которым относится хамса или анчоус. Подробные признаки этой рыбы можно найти в работах А. И. Александрова (1926) и А. А. Майоровой (1934).
Измерение рыб тресковых
(Gadidae)
Треска имеет огромное промысловое значение. Эта рыба распространена в морях и в Атлантическом, Северном Ледовитом и Тихом океанах. Известно, что треска разных мест имеет свои особенности.
Соматические отличия разных групп трески пока изучены неполно, хотя и в этой области мы уже имеем ряд работ (Шмидт, 1904; Дерюгин, 1920; Михин, 1925; Талиев, 1931; Есипов, 1931 и др.), а в 1948 г. вышла монография А. Н. Световидова «Трескообразные».
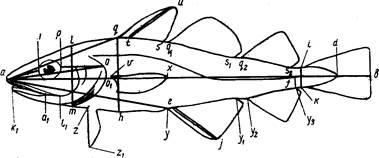
Рис. 11. Схема измерений тресковых рыб (по Михину, 1925 с дополнениями) :
ab-длина всей рыбы; ad-длина без С; o1d-длина туловища; an-длина рыла; пр-диаметр глаза (горизонтальный); ро– заглазничный отдел головы; aо-длина головы; lm-высота головы у затылка; aa1-длина верхней челюсти; k1l1-длина нижней челюсти; qh-наибольшая высота тела; ik-наименьшая высота тела; aq-антедорсальное расстояние; ay-антеанальное расстояние; fd–длина хвостового стебля; qs-длина основания ID; q1s1– длина основания IID; q2s2-длина основания IIID; tu-наибольшая высота ID; уу1-длина основания IA; y2y3-длина основания IIA; ej-наибольшая высота IА; vx-длина Р; zz1-длина V; zy – расстояние между V и IА.
Схема промеров трески (рис. 11) впервые дана в 1925 г. В. С. Михиным. Кроме указанных в этой схеме промеров, измеряются ширина лба, а из меристических признаков определяют число лучей в спинных и анальных плавниках (в каждом отдельно) и число жаберных тычинок.
Величины всех частей тела трески нужно, казалось бы, высчитывать по отношению к длине рыбы без С, придавая последнему термину обычное понимание, т. е. к величине расстояния от конца рыла до конца чешуйного покрова, а не к длине всей рыбы. Индексы В. С. Михин вычислял по отношению к длине всей рыбы, это необходимо иметь в виду.
Рис. 12. Способ определения горбатости трески (по Талиеву, 1931).
Наиболее характерными признаками для различия некоторых групп трески (например, беломорской и мурманской) пока считают высоту тела, высоту головы, ширину лба, длину хвостового стебля, высоту первого спинного и первого анального плавников и число тычинок на первой жаберной дужке. В работе В. С. Михина приведено сравнение многочисленных признаков двух форм трески и дана сводная таблица пределов колебаний признаков той и другой формы.
Д. Н. Талиев (1931) предложил определять угол, составленный прямой, касательной верхней точки тела, продолженной вперед, и линией спины, идущей вперед от ID. Автор дает такое пояснение к последнему признаку: «В работах по систематике рыб, насколько мне известно, измерение углов различных частей тела рыб не производилось, но мне кажется, что введение этих Определений может дать хорошие результаты». Действительно, мы очень часто читаем в систематических работах: «рыба более горбата» и т. п., но все эти внешние признаки (рыбоводы их называют экстерьером рыб) не учитываются точно. По данным Д. Н. Талиева, в двух формах беломорской трески особенно рельефно было видно различие между горбатостью типичной беломорской трески и большей прямизной линии спины у новой формы. Способ измерений трески Д. Н. Талиевым изображен на рис. 12.
При изучении систематики трески принимают во внимание форму и величину плавательного пузыря (Европейцева, 1937 и др.), отношение длины последних лучей 3-го спинного плавника к длине промежутка между D и С (Ильин и Певзнер, 1939).
В 1936 г. вышла работа М. И. Маркуна о налиме р. Камы, где автор на основании измерений сделал описание новой формы налима (Lota lota kamensis). М. И. Маркун брал следующие просчеты и промеры: число лучей в I D, IID, Р, V, А, пилорических придатков, жаберных тычинок, позвонков, длину рыбы без С (с этой длиной Маркун ведет сравнение других длин), наибольшую высоту тела, наименьшую высоту тела, антедорсальное, антевентральное, антеанальное расстояние, длину головы, длину основания I D, длину основания II D, высоту II D, длину основания А, высоту Л, длину Р, длину V, расстояние А-Р (или антепекторальное расстояние, И. П.), расстояние V-А, длину рыла, заглазничный отдел головы, диаметр глаза, высоту головы, ширину лба, длину верхней челюсти, длину нижней челюсти. Последние семь признаков (начиная с длины рыла) выражены в процентах длины головы, остальные – в процентах длины рыбы без С.
Наибольшую высоту рыбы брали у основания I D, наименьшую-по вертикали конца II D и Л (т. е. высота у основания С); высоту II D и Л измеряли по первому лучу, длину V- до конца первого луча этого плавника.
Представляет интерес способ подсчета лучей в П Dи в A. Плавники вырезали и наклеивали на стеклянную пластинку; при’ высыхании перепонок между лучами плавник плотно пристает к стеклу, «и лучи отчетливо выделяются как при проходящем, так и падающем свете», при просчете лучей лучше пользоваться лупой или бинокулярным микроскопом.
Схема измерений налима, разработанная М. И. Маркуном, очевидно, будет принята и другими исследователями. Таким образом, налима можно измерять не по общей схеме измерений тресковых, а по специальной схеме.
А. Н. Световидов в упомянутой монографии «Трескообразные» (1948) при систематическом пазгоаничении многочисленных представителей отряда Gadiformes пользовался схемой В. С. Михина, но брал меньшее количество измерений. Важное систематическое значение А. Н. Световидов придает расположенным на голове порам, ведущим в каналы системы боковой линии.
Б. С. Ильин и В. И. Певзнер (1939) в числе признаков беломорской трески брали, помимо признаков, указанных В. С. Михиным и Д. Н. Талиевым, «большую длину последних лучей спинного плавника по отношению к размеру промежутка между спинным и хвостовым плавниками».
Измерение рыб окуневых (Percidae) и скорпеновых
(Scorpaenidae)
Схема измерения окуня в 1902 г. дана Н. А. Варпаховским в книге «Рыболовство в бассейне р. Оби».
В 1930 г. В. В. Покровский дал предварительную схему измерений окуня, а в 1951 г. после детального исследования морфологических признаков окуней Европы, Азии и Америки предложил новую схему измерений для этого вида ( рис. 13).
После работ В. В. Покровского схему измерений, которую предлагал Варпаховский (1898 и 1902), можно считать излишней.
Длина всей рыбы (аb)-от конца рыла до вертикали от самого длинного луча хвостового плавника (чаще бывает длиннее нижняя лопасть).
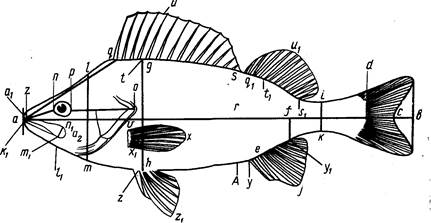
Рис. 13. Схема измерений окуневых (Percidae):
ab-длина всей рыбы; ас-длина по Смитту; ad-длина без С; od- длина туловища; an – длина рыла; пр – диаметр глаза (горизонтальный); ро – заглазничный отдел головы; ао – длина головы; lm – высота головы у затылка; a1a2– длина верхнечелюстной кости; n1m1 – ширина верхнечелюстной кости; k1l1– длина нижней челюсти; gh – наибольшая высота тела; ik – наименьшая высота, тела; aq – антедорсальное расстояние; rd – постдорсальное расстояние; аг- антевентральное расстояние; ау – антеанальное расстояние; fd – длина хвостового стебля; qs – длина основания ID; q1s1 – длина основания IID; tu-наибольшая высота ID; t1u1 – наибольшая высота IID; уу1 – длина основания A; ej – наибольшая высота A; vx – длина Р; vх1 – ширина основания Р; zz1 – длина V, vy – расстояние между Р и А; zу – расстояние между V и A; Ay-расстояние между anus’ом и А.
Длина рыбы без С, т. е. от конца рыла до конца чешуйного покрова (ad). За конец чешуйного покрова принимают самые мелкие последние чешуйки по средней линии тела, покрывающие и хвостовой плавник в его основании.
Чешую в боковой линии надо просчитывать от жаберной щели до последней (включительно) чешуи, имеющей отверстие; за этой чешуйкой имеется еще несколько рядов менее правильно расположенных чешуек, но без боковой линии, при просчете они не принимаются во внимание.
При просчете позвонков уростиль считается за один (последний) позвонок.
Во втором спинном плавнике просчитываются и записываются отдельно колючие лучи (обычно II), мягкие, неветвистые (обычно I) и мягкие ветвистые (от 12 до 15). Пример записи: II, I 13 (колючие лучи от мягких отделяются запятой).
Обычно у окуня имеется по 7 жаберных лучей.
Тычинки на первой жаберной дуге просчитываются отдельно на нижней (длинной) и на верхней (короткой) ветвях. Включать в просчет нужно и зачаточные (рудиментарные) тычинки, имеющие вид малозаметных бугорков. Образец записи: 18+6; 20+7 и т. п.
У жаберной дуги измеряются отдельно обе половины дуги, причем за крайние точки измерений берутся зачаточные тычинки. Пример записи: 20+6=26.
Длину наибольшей жаберной тычинки обычно измеряют у места соединения нижней и верхней ветвей дуги.
Длину головы измеряют от конца рыла до конца postoperculum, а высоту-у затылка. Верхняя точка измерения лежит у начала чешуйного покрова.
Длина рыла-от конца рыла до окрашенной части глаза- радужины.
Диаметр глаза берут горизонтальный – длина окрашенной части глаза.
Длина верхнечелюстной кости измеряется, как у лососевых (без praemaxillare).
Длина нижнечелюстной кости (dentale) – до точки ее сочленения с articulare (в этом месте снаружи заметно небольшое углубление).
Расстояние от ануса до А-от заднего края анального отверстия до начала основания анального плавника.
Хвостовая выемка – отношение линии от наружного края средних лучей хвостового плавника до линии, соединяющей концы верхней и нижней лопастей плавника к этой последней линии. При этом измерении необходимо развернуть хвостовой плавник, придав ему естественное положение.
Работа В. В. Покровского по систематике окуня может служить образцом для таких исследований.
Предложенная В. В. Покровским схема измерения обыкновенного окуня (см. приложение VII) пригодна и для другого 1 представителя того же рода – балхашского окуня (Реrса schrenki), но у последнего при измерении длины тела нужно брать передний край нижней челюсти, потому что она у «его выдается за край верхней челюсти. Схема В. В. Покровского может применяться и при измерении всех других рыб семейства Percidae, для родов: судак (Lucioperca), чопа (Aspro), перкарина (Percarina) и ерш (Acerina). По этой схеме произведено обстоятельное исследование судака (Кудерский, 1958). Этой же схемой с некоторыми изменениями можно пользоваться и для измерений рыб скорпеновых (Scorpaenidae), например, для измерений морского окуня (Sebastes marinus) и для рыб семейства Serranidae, но для морского окуня предложена В. И, Травиным особая схема. В приложении VIII дана ведомость индексов. Кроме того, предлагается обращать внимание на характер исчерченности и форму operculum и других костей черепа.
Измерение рыб атериновых
(Atherinidae)
Рыбы, относящиеся к этому семейству, представляют интерес в систематическом отношении, и притом они мало изучены. Г. И. Венеров, занимавшийся изучением признаков каспийской атерины (Atherima mochon pontica n. caspia), рекомендовал давать следующие промеры тела этой рыбы: длина всей рыбы, длина тела до конца средних лучей С, длина тела без С, длина туловища, количество поперечных рядов чешуи (squamae), так как боковой линии у атерин нет, количество лучей в плавниках, количество позвонков, количество жаберных тычинок, длина рыла, диаметр глаза, заглазничный отдел головы, длина головы, высота головы у затылка, высота головы через середину глаза, ширина лба, длина верхнечелюстной кости, ширина верхнечелюстной кости, длина нижней челюсти, длина первой жаберной дужки, длина наибольшей жаберной тычинки, наибольшая высота тела, наименьшая высота тела, наибольшая толщина тела, антедорсальное расстояние, постдорсальное расстояние, антевентральное расстояние, антеанальное расстояние, длина хвостового стебля, расстояние Р-V, расстояние V-А, длина основания ID, высота ID, длина основания А, высота А, длина Р, длина V, длина основания IID, высота IID, расстояние между ID и IID, величина чешуи, ширина серебристой полоски тела, количество рядов зубов на челюстях.
Измерение рыб кефалевых
(Mugilidae)
А. М. Попов в статье о кефалях (1930) приводит большое количество описательных признаков и прилагает список 73 признаков кефалей, который им назван схемой для измерения рыб семейства Mugilidae.
К сожалению, автор не дал необходимых пояснений к этим признакам и к определениям индексов, а также допустил в тексте статьи, в ведомостях промеров и в схеме измерений досадные описки, указывая число чешуи в боковой линии. У кефалей нет боковой линии, понимаемой в общепринятом ихтиологическом смысле. Поэтому под количеством чешуи в боковой линии надо понимать количество поперечных рядов чешуи (по боку тела), т. е. squamae.
Л. С. Берг (1949) в сводке систематических признаков 6 видов кефалей рода Mugil, встречающихся в водах СССР, ограничился краткой определительной таблицей, составленной главным образом на таких признаках, как размеры подглазничной кости, на особенностях жирового века, на толщине верхней губы, на характере чешуйных канальцев, на количестве чешуи от конца рыла до первого спинного плавника, на присутствии или отсутствии удлиненной лопастинки над основанием брюшного плавника. Таблица Л. С. Берга позволяет распознавать каждый из шести видов кефалей. При изучении внутривидовых разностей кефалей, несомненно, потребуется введение большого числа признаков, вернее, потребуется большое количество промеров и просчетов, но все же едва ли так много, как указано в схеме А. М. Попова. В. В. Абрамов (1952), указывая на случаи нахождения лобана (Mugil cephalus) в низовьях Амура, приводит пластические и меристические признаки одного экземпляра этой рыбы, пользуясь в основном схемой измерений карповых рыб с добавлением следующих признаков: длина тела до конца средних лучей С, ширина щели в веке, высота верхней губы, длина удлиненной чешуйки над Р, длина такой же чешуйки над V, число жаберных лучей, жаберных тычинок и количество «продольных серых полос по левому боку тела», длина основания и высота второго D. Диаметры глаза приняты во внимание оба (горизонтальный и вертикальный). Автор подметил, что «по середине базального края чешуи имеется выемка».
Измерение рыб песчанновых
(Ammodytidae)
Полового диморфизма у песчанки (Ammodytes) не подмечено, поэтому можно пользоваться, по совету Е. К. Суворова, Л. А. Вадовой и А. Н. Сынковой (1931), смешанным материалом.
В работе В. И. Певзнера (1928) имеется указание, которое; своей категоричностью может создать ошибочное представление о расовом составе песчанок Баренцева моря. Автор проанализировав всего лишь три меристических признака (число позвонков, лучей в D и лучей в Л) песчанок, доставленных из Териберки и Восточной Лицы, пришел к выводу, что «на этом основании мы не можем говорить о существовании различных местных групп песчанки, так как пробы из Териберки вполне идентичны с пробой из Восточной Лицы».
В 1937 г. вышла работа по систематике песчанок Г. У. Линдберга. В этой работе обосновывается существование в водах северного полушария трех видов песчанок.
Измерение рыб семейства
Cobitidae [гольцы (Nemachilus). губачм
(Diplophysa), вьюны (Misgurnus),
щиповка (Cobitis)]
Разработанной схемы промеров рыб этого семейства нет. В значительной мере пригодна для этой цели схема промеров карповых, но к ней необходимы значительные дополнения. Тело у некоторых представителей семейства Cobitidae голое (в ведомости промеров в таких случаях в графе, относящейся к боковой линии, надо ставить 0). Число усиков разное, следовательно, нужно указывать количество усиков и их длину. Нужно указывать (описательно) форму хвостового плавника (усеченный, с выемкой, округлый), наименьшую высоту тела выражать в процентах длины хвостового стебля; у вьюнов просчитывать число поперечных рядов чешуи (squamae), указывать число темных пятен на боках. Для всех рыб этого семейства нужно давать (на 1-4-й страницах таблицы промеров) описание окраски тела.
Рыбы семейства Cobitidae в морфологическом и вообще в систематическом отношении представляют много особенностей, начиная со стадии личиночного развития. А. В. Гриб (1936). изучая личинок вьюна (Misgurnus fossilis), обнаружил, что они имеют наружные жабры. Изучение взрослых рыб рода Misgurnus надо вести отдельно для самцов и самок, так как в некоторых признаках у них выражен половой диморфизм (у самцов второй луч Р удлинен).
Большие трудности представляет изучение признаков рыб рода Nemachilus, в который входит большое количество видов. Видовые признаки до сих пор еще недостаточно определены, вследствие чего Л. С. Берг (1948) говорит: «Разбить этот род на основании до сих пор предложенных признаков невозможно». Помимо промеров, приходится описывать форму хвостового плавника (с выемкой или без выемки), пятна и полосы, имеющиеся на теле, и указывать, имеется ли чешуя.
При изучении губачей нужно измерять не только длину, но и толщину хвостового стебля (и сравнивать толщину хвостового стебля с его высотой, т. е. с наименьшей высотой тела). Схема промеров губача Nemachilus strauchi из Балхаша дана В. В. Покровским и К. И. Беляевой (1948).
У щиповки (род Cobitis) нужно определять простой или двухраздельный подглазничный шипик, а также указать толщину тела. Существенное значение в систематической группировке щиповок имеет и характер окраски тела.
Д. Н. Талиев (1946) установил новый вид бычка из оз. Баунт на основании изучения следующих признаков: числа лучей в D, А, Р, V, С, количества чешуи в l. l., наибольшей и наименьшей высоты тела, длины хвостового стебля, длин основания I и II D, основания А, длины Р и V, антедорсального расстояния, длины головы, заглазничного пространства, диаметра глаза, ширины лба, длины рыла, высоты головы, ширины головы, ширины лба. Принимались также во внимание строение зубов, размер жаберного промежутка, кожные шипики и бугорки. А в другой работе (Талиев, 1946) тем же автором дается диагноз также нового вида бычков (Metacottus gurwici) на основании особенностей строения скелета (степень развитости postcleithrum, характер парапофизов туловищных позвонков и др.).
Измерение рыб семейства
(Gobiidae)
Для бычков семейства Gobiidae нет опубликованной схемы промеров, но по специальным работам такую схему нетрудно предложить. В статье П. О. Зубовича (1925) опубликовано описание только одной формы черноморского бычка. Автор дает перечень следующих признаков: длину всей рыбы от начала головы до конца хвостового плавника; длину тела без хвостового плавника, от начала головы до начала хвостового плавника, т. е. до того места, где имеется перегиб; длину головы;
ширину головы (измерение производилось в плоскости бороздок, отделяющих щеки от жаберных крышек); высоту головы (в том же месте, где определялась ширина головы); межглазничный промежуток или ширину лба (в самом узком месте); диаметр глаза (у бычков глаз круглый); заглазничное пространство; длину рыла; наибольшую высоту тела (в плоскости третьего луча первого спинного плавника); наименьшую высоту тела (в самом низком месте хвостового стебля); наибольшую ширину тела (в том месте, где бралась наибольшая высота тела); наименьшую ширину тела (в плоскости конца анального плавника); длину хвостового стебля; расстояние от начала головы до анального отверстия; расстояние от конца хвоста до анального отверстия; длину хвостового плавника; длину Р; длину V ==(Р и V измерялись приложенными к телу); длину основания II D; длину основания анального плавника; расстояние V-A; антедорсальное расстояние; антеанальное расстояние; длину луча I D; длину луча II D; число лучей в II D; число лучей в А; число рядов чешуи. Помимо того, автор дает определение пола и степени зрелости половых продуктов (по пятибалльной системе) и вес в граммах.
В работе Зубовича описаны две категории присасывательных дисков, свойственных двум разным видам бычков, а также приводятся промеры длины начальных, средних и конечных лучей D и А.
При изучении систематической группировки бычков этого семейства решающее значение имеет описание так называемых пор и слизистых каналов, расположенных на голове бычков. На это обращено большое внимание многими авторами. Среди советских исследователей с особенным успехом работал в этом направлении Б. С. Ильин. Томияма (1936) описывает более 100 форм бычков из Японии, пользуясь довольно простой схемой измерений, опубликованной в его работе. Главными отличительными признаками для рыб семейства Gobiidae он избрал длину тела без С, длину головы, диаметр глаза, наибольшую И наименьшую высоту тела, а также число рядов чешуи. Автором дана и схема промеров.
Измерение рыб угревых
(Anguillidae)
Выдающиеся работы, проведенные датским исследователем И. Шмидтом (Ioh. Schmidt, 1906; 1923), осветили многие явления из жизни угрей, считавшиеся ранее загадочными. Имеются и новые наблюдения. –
Главнейшим признаком, характеризующим отдельные виды угрей [европейский (речной), американский и японский угри] Шмидт считает количество позвонков; существенно определять также количество лучей в плавниках, особенно в анальном (просчитывают число лучей и в других плавниках, т. е. в D, Р и В С, брюшных плавников угри не имеют). Нужно определять число жаберных лучей (жаберных тычинок речной угорь не имеет) и число отверстий в боковой линии. Из пластических признаков нужны: длина рыла (от переднего края нижней челюсти при закрытом рте), диаметр глаза, ширина лба, длина головы, высота головы по середине глаза, наибольший обхват тела, наибольшая толщина тела, наибольшая высота тела, антедорсальное расстояние, антеанальное расстояние, расстояние от конца рыла до анального отверстия и размеры плавников. Кроме того, следует отмечать окраску тела. У неполовозрелых угрей брюшко желтовато-белое, у половозрелых-серебристо-белое. За длину рыбы, с которой сравнивают части тела, принимают всю или абсолютную длину (от конца рыла до конца хвостового плавника).
В ведомости индексов отдельные части тела угря нужно высчитывать по отношению к абсолютной длине тела, а малые величины (рыло, глаз, ширина лба, высота головы) – к длине головы. Диаметр глаза следует высчитывать в процентах длины рыла.
Исключительный интерес приобретает вопрос изучения молоди угря. В наши пресные воды входит угорь в молодом возрасте (3 лет). В пресноводных водоемах угри живут от 4 до 7 лет (почти одни самки). За это время угорь растет и становится половозрелым. В книге Эренбаума (Ehrenbaum, 1930) сделана подробная сводка всех сведений по систематике и биологии речного угря. О распространении угря по водоемам СССР есть сводка Л. С. Берга (1916), по биологии угря из Финского залива-работа В. С. Михина (1939).
В настоящее время в Белоруссии ведутся работы по выращиванию угря в озерах и опубликовано несколько ценных работ С. В. Кохненко (1954, 1958), С. В. Кохненко и Е. А. Боровик (1957) по биологии и по морфометрии молоди и взрослых угрей. В статье «Морфологическая характеристика стекловидных угорьков» С. В. Кохненко и Е. А. Боровик описывают и методику исследования признаков угрей этой стадии развития. Промеры угорьков производились циркулем-измерителем, кронциркулем и окуляр-микрометром. Они определяли: длину всего тела, антедорсальное и антеанальное расстояния, расстояние от переднего края рыла до ануса, длину спинного, грудного, хвостового и анального плавников, наибольшую высоту и толщину тела, расстояние от D до Л, длину, высоту и ширину головы, длину рыла, ширину лба, диаметр глаза. Просчитывали позвонки и брали отолиты. Получены интересные данные, показывающие, что соотношения морфологических признаков с возрастом рыбы изменяются иначе, чем у других рыб. Так, относительная длина головы угрей «с возрастом изменяется мало», высота и ширина головы с возрастом изменяются «в сторону уменьшения, а ширина лба – в сторону увеличения». Диаметр глаза в течение пресноводного периода жизни угря слабо изменяется.
Для измерения речного угря опубликована схема С. В. Кохненко (1958). В основу схемы положены промеры карповых, но включены многие дополнительные признаки: обхват головы по середине глаз, длины пищеварительного тракта, пищевода, желудка, кишечника, расстояния между передними носовыми отверстиями и между задними носовыми отверстиями. Индексы промеров определены по отношению к длине всего тела, а промеры частей головы, кроме того, выражены в процентах длины головы. Из меристических признаков взяты: число позвонков, отверстий в боковой линии, лучей в Р, D, С и А и число жаберных лучей.
У взрослого речного угря полностью отсутствуют жаберные тычинки (рис. 14).

Рис. 14. Жаберный аппарат взрослого угря – полное отсутствие жаберных тычинок.
Интересно изучить структуру жаберного аппарата на разных стадиях развития угря, начиная с личиночного периода. Мой просмотр жаберных дужек стекловидных угорьков и угрей, пробывших в пресном водоеме одного год, указывает на присутствие на первой дужке бугроватых неровностей, которые как бы соответствуют зачаточным тычинкам. Но эти наблюдение следовало бы продолжить.
Измерение рыб камбаловых
(Pleuronectidae)
Отряд камбалообразных (Pleuronectiformes) рыб имеет не только много родов и видов, но и несколько семейств. Однако все представители этой группы морфологически должны быть признаны сходными, и для них следует иметь одну общепринятую схему измерений. Многочисленные исследователи, занимавшиеся изучением расового состава камбаловых, установили большое расовое разнообразие указанных рыб.
Дункер (Duncker, 1896) предложил схему измерений камбал и в нескольких статьях дал описание различных форм камбал, пользуясь своей схемой. В последующих работах по камбаловым рыбам схема Дункера претерпела некоторые изменения.
В 1934 г. вышла монография Нормэна (Normann, 1934) по систематике камбаловых, где есть данные о различительных признаках камбалы.
Е. К. Суворов (1929) привел схему измерений для камбалы Чешской губы и по этой схеме выполнил биометрическую работу по описанию двух новых форм камбал (Pleuronectes flesus caninensis и Pl. glacialis caninensis), причем на основании вариационно-статистического анализа сделал оценку признаков. Следует заметить, что автор располагал очень малым в количественном отношении материалом, лишь 13-15 особей одной группы и 17 особей другой группы (такой материал вариационно-статистическим методом обрабатывать не рекомендуется).
В 1930 г. вышли ценные работы по систематике камбал Баренцева и Белого морей Н. В. Сыч-Аверинцевой (1930). В этих работах можно найти много полезных методических указаний как по счетным, так и по измерительным признакам.
Изучением камбаловых (палтусов) занималась в 1935 и 1936 г. М. Ф. Вернидуб, использовавшая опыт предшественников и давшая свои оригинальные указания. В своей работе в 1935 г. она дала схему измерения палтусов (рис. 15), в основу которой положена схема измерений камбал рода Pleuronectes, предложенная Дункером. М. Ф. Вернидуб ввела новое понимание признака высоты головы, считая таковой расстояние по вертикальной линии, идущей, касаясь заднего края глазницы, наиболее удаленного (от рыла) края глаза. Едва ли есть основание принимать этот признак в понимании М. Ф. Вернидуб, так как высота головы камбалы слишком условна.
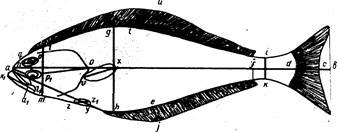
Рис. 15. Схема измерений палтусов (по Вернидуб, 1936):
ab-длина всей рыбы; ас-длина до конца средних лучей С; ad-длина без С; an-длина рыла; пр-диаметр глаза (горизонтальный); р1о- заглазничный отдел головы; ао – длина головы; lm – высота головы; аа1–длина верхней челюсти; k1l1–длина нижней челюсти; gh-наибольшая высота тела; ik – наименьшая высота тела; aq – антедорсальное расстояние; аz-антевентральное расстояние; ay-антеанальное расстояние;, fd-длина хвостового стебля; tu-наибольшая высота D; ej-наибольшая высота A; vx- длина Р; zz1 – длина V; zу – расстояние между V и A; dc – длина средних лучей С. Измеряется также ширина лба.
Из счетных признаков для палтусов М. Ф. Вернидуб берет: число чешуи в l. l., число лучей в D, А и Р, числа тычинок на первой жаберной дужке, туловищных позвонков и хвостовых позвонков. Из пластических признаков М. Ф. Вернидуб брала (в процентах длины тела до конца средних лучей С): длину тела до С, длину туловища, хвостового стебля, головы, наибольшую и наименьшую высоту тела, высоту головы, антедорсальное, антеанальное и антевентральное расстояния, расстояние V-А, длину Р на зрячей стороне, длину Р на слепой стороне, высоту D, высоту А, длину верхней и нижней лопастей С, длину средних лучей С. Помимо этого высчитаны в процентах длины головы, заглазничный отдел головы, длина рыла, длина верхней челюсти зрячей и слепой стороны, горизонтальный диаметр нижнего и верхнего глаз, вертикальный диаметр нижнего и верхнего глаз и межглазничное пространство. Наконец, М. Ф. Вернидуб определяла в процентах длины тела длину и высоту хвостового стебля.
При изучении камбаловых, особенно палтусов, необходимо обращать внимание на расположение и количество нижнеглоточных зубов, а также зубов нижней и верхней челюстей, и давать описание формы чешуи. Помимо промеров, следует давать описание плавников (брюшные плавники симметричны или асимметричны), указывать, на какой стороне глаза, процент левоглазных и правоглазных, прямая или изогнутая боковая линия, есть или отсутствуют костяные бугорки у оснований D и А, окраска тела, количество и характер пилорических придатков, характер нижнечелюстных зубов. На эти признаки обращают внимание Нормэн (1934) и Г. У. Линдберг с Д. И. Охрямкиным (1936).
В области изучения систематики камбаловых предстоит сделать еще многое. Прежде всего следует разрешить вопрос о стандартной длине тела, по которой нужно вести сравнение частей тела: одни такой длиной считают длину тела без С (Суворов, Вернидуб), другие-длину всей рыбы (Сыч-Аверинцева). Так как нередко количество лучей в левом и правом грудном плавнике различно, следует вести счет лучей в левом и правом Р.
Много методических указаний содержит книга П. А. Моисеева «Треска и камбалы дальневосточных морей» (1953).
Измерение миноговых
(Petromyzonidae)
Для миног, имеющих весьма отличные от настоящих рыб морфологические признаки, рекомендуем пользоваться специальной схемой, приведенной на рис. 16 и в приложении IX. К этой схеме необходимы следующие пояснения. Вся длина у миног совпадает с длиной тела, потому что задняя часть хвостового плавника лишь незначительно (на 1-2 мм) увеличивает общую длину. Для определения диаметра глаза измеряется просвечивающее сквозь кожу глазное яблоко. Высота спинных плавников определяется размерами наиболее длинных их лучей по наклонной линии. Обязательно определение количества и характера зубов на челюстных и язычной пластинках: это один из главнейших систематических признаков миног. Ротовая воронка миноги изображена на рис. 17. Способа определения возраста миног пока не найдено.
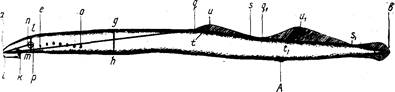
Рис. 16. Схема измерений миноговых (Petromyzonidae):
ab-вся длина; gh-наибольшая высота тела; lm-высота головы; ае – расстояние от конца рыла до первого жаберного отверстия; an – длина рыла; пр-диаметр глаза (горизонтальный); ре-промежуток между глазом и жаберным аппаратом; ik-диаметр ротового диска; ао – длина головы с жаберным аппаратом; аq- антедорсальное расстояние; аА – расстояние от конца рыла до ануса; sq1 – промежуток между I D и II D; qs – длина основания I D; tu – наибольшая высота I D; q1s1.-длина основания II D; t1u1–наибольшая высота II D; s1b-длина спинной части С; Аb-расстояние от ануса до конца С.
|
|
Рис. 17. Ротовая воронка миноги из рода Lampetra (по Бергу, 1948):
Верх. г. з.- верхнегубные зубы; В.- ч. пл.- верхнечелюстная пластинка; Вн. бок. г. з.- внутренние боковые губные зубы; Н.-ч. пл.-нижнечелюстная пластинка; Краев, з.- краевые зубы; Ниж. г. з.-нижнегубные зубы; Пер. яз. пл.- передняя язычная пластинка; Внеш. бок. р.. з.- внешние боковые губные зубы; Рот. отв.- ротовое отверстие.
Так как плавники миног не имеют косточек, поддерживающих плавниковую складку, то высоту плавников можно правильно определить лишь на экземплярах сильно влажных. Для удобства промеров плавничок можно приколоть к столу булавками, да и самую миногу при промерах расправлять на столе также лучше булавками.
Благодаря работам Л. С. Берга и М. М. Ивановой-Берг (1936) установлено, что у речной миноги (Lampetra fluviatilis) существуют две расы, которые идут на нерест (в реку Неву) в разные сроки: одна – весной, другая – осенью, причем морфологических различий между этими биологическими группами пока не установлено.
Следует заняться установлением признаков личинок миног.
Замечания по измерению других рыб
Для многих семейств рыб еще нет установленных схем промеров. Но современная ихтиологическая литература все более, все чаще обогащается и новыми схемами измерений рыб и списками многочисленных признаков рыб. Например, в книге Фаулера (Fowler, 1936) приведена подробная схема измерений рыб, которой он пользовался при описании морских рыб Западной Африки. Схема (для разных видов рыб) имеет около-40 наименований частей тела, из которых названы: длина головы, наибольшая высота тела, длина хвостового стебля (кстати сказать, измеряемая Фаулером иначе, чем принято), предорсальное расстояние (измеряемое также иначе), длина рыла,. диаметр глаза, части верхней и нижней челюстей, расстояние между ноздрями, просчитывается число чешуи в l. l. над l. l. и под l. l., длина всей рыбы, длина тела без С и т. д.
Ниже мы даем некоторые замечания, которые могут оказать пользу лицам, приступающим к описанию морфологических признаков рыб, еще не имеющих определенных схем промеров.
Акулы (группа Selachoidei). Признаки: количество жаберных щелей, длина всей рыбы (к этой длине относят промеры других частей тела), наибольшая высота тела, длина головы до первой жаберной щели, толщина головы, диаметр глаза, межглазничное пространство, длина рыла, расстояние от конца рыла до I D, антевентральное расстояние, длина основания II D, высота ID (от скрытого в коже основания самого длинного луча), высота II D, высота колючки I D (если она есть), высота колючки II D, длина Р, длина V, расстояние между ноздрями. Фаулер при описании акул обращает внимание на форму рта, его положение и вооруженность зубами. Нужно описывать имеющиеся на теле шипики. Л. С. Берг (1911) при классификации акул обращает внимание -на хвостовые кили, вырезку С, зубы, размеры жаберных щелей, расположение I D, на положение spiraculum и ноздрей.
Скаты (группа Batoidei). Длина диска от верхушки рыла до середины линии, соединяющей задние края грудных плавников, число и характер шипов по средней линии тела, длина всего тела (хвостовой бич, если он есть, измерять отдельно), наибольшая ширина тела, длина рыла, ширина лба, диаметр рта, расстояние от рта до вершины рыла, диаметр глаза, диаметр spiraculum, расстояние от ноздри до вершины рыла, длина I D, длина II D, расстояние между I и II D, длина головы, расстояние между ноздрями и длина хвоста. Л. С. Берг указывает, что у скатов длина рыла с возрастом сильно увеличивается. Вообще многие признаки скатов имеют возрастную изменчивость (колючки плавников, хвостовые шипы, форма зубов, окраска и др.).
Щуковые (Esocidae). Длина рыбы без С (длина тела измеряется от передней точки нижней челюсти), длина туловища, количество чешуи в боковой линии, над ней и под ней (вниз до основания Л), число поперечных рядов чешуи (нужно учитывать, что боковая линия у молодых щук обычно незаметна или неполная, у взрослых полная, squamae, число чешуи по боку хвостового стебля, число лучей в D, P и V (просчет лучей в Р и V необходим, потому что этот признак у щук обнаруживает значительные колебания) и в Л, числа тычинок на первой жаберной дуге (тычинки у взрослых щук в виде бугорков), жаберных лучей, позвонков, длина рыла, диаметр глаза, заглазничный отдел головы, длина верхнечелюстной кости, длина нижней челюсти, ширина лба, длина головы (от переднего края нижней челюсти), высота головы у затылка, высота головы через середину глаза, наибольший обхват тела, наибольшие толщина и высота тела, наименьшая высота тела, длина хвостового стебля, антедорсальное, антевентральное, антеанальное расстояния, длина D, высота D, длина Р, высота V, длина Л, высота Л, расстояние между P-V, расстояние V-A, длина верхней лопасти С; длина нижней лопасти С и длина средних лучей С.
А. И. Ефимова (1949), описывая щуку Обь-Иртышского бассейна, установила, что имеются большие колебания в количестве чешуи в боковой линии, а также в числе лучей в D и Л. Она показала, что у самцов щуки Р и V длиннее, чем у самок. При описании щук нужно обращать внимание на окраску тела и на расположение пятен.
Сомовые (Siluridae). Ф. Н. Бизяев выполнил большую работу по установлению морфологических признаков сома из дельты р. Дона и Таганрогского залива, применив свою схему измерений. Исследователь брал очень много промеров и просчетов. Так как подобное изучение этого вида рыб проводится впервые, естественно, нужно проанализировать возможно большее количество признаков, чтобы облегчить дальнейшую такую же работу над сомами из других мест. Автор принимал такие признаки:
число лучей в D, Р, V, А и С, число трубочек по боку тела, позвонков во всем позвоночнике, прекаудальных и каудальных позвонков, жаберных тычинок и жаберных лучей. Из других признаков брал длину и высоту головы у затылка и через середину глаза, длину верхней и нижней челюстей, предзаглазничное и заглазничное пространства, диаметр глаза, ширину лба, длину виска, усиков, наибольшую высоту тела, высоту тела около анального плавника, длину основания и высоту D, А, Р, V, измерял ширину и длину пасти, наибольший обхват тела, обхват тела у анального плавника, длину горла, антедорсальное, постдорсальное, антепекторальное, пектовентральное, антеанальное, анально-каудальное, антевентральное и поствентральное расстояния, расстояние между V и Л. Пластические признаки высчитывал в % всей длины тела; устанавливал длину тела до основания хвостового плавника и длину туловища. В % длины головы выражал все части головы, включая длину усиков и длину горла («пространства от места прикрепления жаберной перепонки до вертикали оснований грудных плавников»).
Рыб, относящихся к группе Jugularis, куда включаются семейства: Trachinidae(рыбы-драконы), Uranoscopidae(звездочеты), Gobiesocidae, Blenniidae, Zoarcidae (бельдюги), Ophi-diidae, Anarhichadidae (зубатки) и др., можно измерять, пользуясь схемой измерений тресковых, в частности по схеме измерений налима, но лучше взять особую схему, например, ту, которую применил Г. У. Линдберг (1938) при изучении некоторых родов и видов семейства Blenniidae. В ведомость измерений автор включил: всю длину тела, длину тела без С, длину головы, наибольшую высоту тела, антедорсальное и антеанальное расстояния, наименьшую высоту тела, длину грудного плавника, горизонтальный диаметр глаза, длину рыла, длину верхнечелюстной кости, межглазничное пространство, длину хвостового стебля, число лучей в D и А. Помимо того, дан целый ряд других признаков: жаберные перепонки прикреплены или не прикреплены к межжаберному промежутку, есть или отсутствует боковая линия и дополнительная линия пор, если на голове есть гребень, то замечается, хорошо ли он развит, покрыто ли тело чешуей, есть ли нёбные зубы и усики над глазами.
Советские ихтиологические исследования, проводимые в океанах, вызвали необходимость разработки новых схем промеров некоторых морских рыб. В обширной статье Н. В. Парина (1960) приведена подробная схема для летучих рыб.
Есть новая схема для лампаниктов (Lampanyctus) и светящихся анчоусов, которыми занималась Е. Б. Куликова (1957). Особенно важна схема расположения фотофоров этих рыб.
Из сказанного о схемах измерений рыб следует, что при первоначальном изучении вида надо пользоваться наиболее подробной схемой, чтобы узнать степень изменчивости каждого признака, в дальнейшем, при исследовании внутривидовых разностей, в схеме оставляют только те просчеты и промеры, которые могут характеризовать ту или другую разновидность внутривидового порядка. При составлении ведомостей индексов показатели малых частей тела (длина рыла, диаметр глаза, ширина лба) лучше вычислять в % длины головы, другие показатели-в % длины тела. При расовых работах также нужны отношения между отдельными частями тела: длина глаза в % длины рыла, наименьшая высота тела в % длины нижней челюсти (у сигов), высота рыльной площадки в % ширины той же площадки (у сигов), длина наибольшей жаберной тычинки в % длины жаберной дужки, ширина верхнечелюстной кости, длина V в % длины Р и др.
Значение в систематике рыб анатомических признаков
Отводя внимание морфологическим признакам, ихтиологи не забывают, что для распознавания многих рыб могут служить и анатомические признаки: количество позвонков, строение ребер, плавательный пузырь, пилорические придатки, вооруженность зубами, размеры икринок и т. п. Много и других подобных примеров (серологический анализ различия рас и т. п.). Но все же морфологический метод изучения рыб по достоинству занимает первенствующее место в систематике рыб как и вообще животных и растений, пользование им и усовершенствование его совершенно необходимо.
Анатомические признаки рыб заслуживают большого внимания. По количеству позвонков различаются расы сельдей (Heincke, Рабинерсон, Аверинцев и др.), угрей (Schmidt, Ehrenbaum), камбал (Duncker, Сыч-Аверинцева и др.). При просчете позвонков должно быть полное однообразие; если препарируется весь позвоночник, то следует при нем оставлять и основную затылочную кость (basioccipitale) и последний хвостовой позвонок.
Препарирование позвоночника не сложно: срезать мускулы с боков, со спины и брюшка, затем позвоночник с неотделенной головой и оставшимися кусками мяса вскипятить (но не переваривать), после чего кости позвоночника очистить от мяса жесткой зубной щеткой.
При массовом просчете позвонков пользуются и рентгеноскопическими приборами, которые освобождают исследователя от препарирования позвоночника.
Число и характер жаберных тычинок рыб в последнее время все более и более привлекают внимание к себе систематиков.
Препарирование жаберного аппарата требует хорошего навыка: надрезать нижнюю часть жаберного аппарата (удобно это делать ножницами), так чтобы при ней остался язык, затем скальпелем отрезать жаберный аппарат в месте соединения глотки с пищеводом, после чего легко отделить верхнюю часть жаберного аппарата. Вынутый через жаберные отверстия аппарат разрезают затем по дорсальной (спинной) стороне.
В работе Б. С. Лукаша (1938) о рыбах реки Вятки указывается, что каспийскую миногу (Caspiomyzon wagneri) легко отличить от миноги речной (Lampetra fluviatilis) не только по характеру зубов на ротовых пластинках, но и по количеству глоточных тычинок: у каспийской миноги таких тычинок 3, у речной миноги 5.
При систематических работах по рыбам нужно просчитывать к описывать зубы – глоточные, челюстные, нёбные, сошниковые.
Форма и вооружение сошника (vomer) у некоторых рыб считается хорошим систематическим признаком. По этому признаку различают Salmo salar и Salmo trutta, беломорскую и мурманскую сельдь (Clupea harengus pallasi maris-albi и Cl. harengus harengus). А. И. Рабинерсон (1923) отметил, что у беломорской сельди сошниковые зубы слабые, у мурманской- сравнительно сильные. Вообще работы по остеологическим признакам рыб представляют большой интерес, но требуют много времени и значительной подготовки. Таких исследований, нужно заметить, у нас немного.
На основании остеологического изучения рыб семейства лососевых хариусы выделены (Чернавин, 1923) в самостоятельное семейство Thymallidae.
Занимающийся систематикой рыб должен знать строение головных костей, из которых при систематических, работах наиболее необходимо отличать: сошник (vomer), этмоидальную (ethmoideum), основную затылочную (basioccipitale), квадратную (quadratum), нёбную (palatinum), предкрышечную (ргаеoperculum), крышечную (operculurn), подкрышечную (suboper-culum), межкрышечную (interoperculum), нижнечелюетную (dentale), верхнечелюстную (maxillare), межчелюстную (inter-maxillare), язычную (glossohyale) кости, жаберные лучи (radii branchiostegi), предглазничные. (praeorbitale), подглазничные (suborbitalia) и носовую (nasale) кости (рис. 18).
Много остеологических признаков приводится в упомянутых уже книгах Л. С. Берга-Система рыбообразных и рыб, А. Н, Световидова-по трескообразным и сельдевым, В, В. Барсукова – по зубаткам, Д. Н. Талиева – по байкальским бычкам.
Нередко приходится просчитывать число пилорических придатков (например, для рыб рода Oncorhynchus). Этот счетный признак сильно колеблется, и считать пилорические придатки трудно. Рекомендуем до просчета выдерживать пищеварительные органы рыбы в спирте (70°); если же они были законсервированы в формалине, то до просчета их следует выдержать в холодной воде (до 24 ч). Самый просчет более удобно вести путем отрывания (пинцетом) каждого отростка.
Рис. 18. Кости головы окуня (по Юдкину, 1951):
I-носовая (nasale); 2-этмоидная (ethmoideum); 3-боковая этмоидная или переднелобная (ethmoideum laterale или praefrontale); 4- лобная (frontale)- 5 – предглазничная (praeorbitale); 6 – подглазничные (subor-bitalia)- 7-основная клиновидная (basisphenoideum); 8-алисфеноид или крылоклиновидная (alisphenoideum); 9- теменная (parietale), 10-клино-видноушная (sphenoticum); 11- переднеушная (prooticum), 12-заднеушная (opistoticum); 13 – крыловидноушная (pteroticum); 14-верхнеушная (epioticum), 15- верхнезатылочная (supraoccipitale), 16- боковая затылочная (occipitale laterale); 17- основная затылочная (basioccipitale); 18- сошник (vomer); 19- парасфеноид (parasphenoideum); 20-межчелюстная или предчелюстная (praemaxillare); 21 – верхнечелюстная (maxillare)- 22- нёбная (palatinum); 23 – внешнекрыловидная (ectopterygoideum). 24 – внутреннекрыловидная (endopterygoideum); 25 – заднекрыловидная (metapterygoideum); 26 – подвисочная (hyomandibulare); 27- соединительная (symplecticum); 28- крышечная (operculum), 29- предкрышечная (praeoperculum); 30 – подкрышечная (suboperculum); 31 – межкрышечная (interoperculum); 32 – нижнечелюстная или зубная (dentale) 33 – сочленовая (articulare); 34-угловая (angulare); 35-квадратная (quadratum); 36-язычная (glossohyale); 37 – нижнеподъязычная (hypohyale); 38 – среднеподъязычная (ceratohyale); 39 – верхнеподъязычная (epihyale); 40 – палочковидная (stylohyale); 41 – лучи жаберной перепонки (radii branchiostegi).
Форму плавательного пузыря и отношение передней и задней его частей (у тех рыб, у которых пузырь состоит из двух мешков, например у карповых) мы относим тоже к числу систематических признаков (у воблы задний конец пузыря тупой, у кутума – острый). Форма и длина отростков плавательного пузыря трески – хорошие признаки, отличающие атлантическую и тихоокеанскую треску.
Структура чешуи тоже является систематическим признаком, но пока по чешуе можно определять только крупные систематические группы-семейства, род (не всегда), редко вид и лишь в исключительных случаях подвид. Между тем, например, при работах по питанию рыб, чешуя часто является единственным признаком, по которому можно определить систематическое положение жертвы (в кишечнике хищников чешуя захваченных ими рыб сохраняется долго в совершенно неизменном виде).
Чешуя различается по своей общей форме, по форме переднего и заднего края, но не менее характерным признаком служат так называемые радиальные линии, идущие от середины (от nucleus) чешуи к ее заднему краю. Число и расположение таких главных линий особенно легко наблюдать даже невооруженным глазом на чешуйках, лежащих близ боковой линии. Так, у типичной плотвы (Rutilus rutilus) 3-5 радиальных линий резко выделяются на общем фоне испещренности чешуи; у серушки (R. rutilus fluviatilis) также несколько таких линий выделяются над другими, но уже не столь резко; у воблы (R. rutilus caspicus) это разнообразие радиальных линий еще более слабо, кроме того, у некоторых особей воблы наблюдается многочисленная испещренность чешуи. У кутума (R. frisii kutum) – подвида довольно близкого к вобле – радиальных линий до двух десятков и резкого выделения одних линий над другими уже почти не заметно. Еще больше радиальных линий у типичного вырезуба (R. frisii); у подуста (Clondrostoma nasus) 6-8 резких радиалей (Kychlicki, 1931). На чешуе леща радиалей много.
В старой немецкой литературе (Heckel und Kner, 1858 и др.) рисунку чешуи придавалось существенное значение.
Следует сказать, что изучение формы чешуи, равно как и формы костей рыб, дает много полезного не только в области распознавания рыб, но и вводит в область филогенетического изучения рыб и в область прошлого ихтиофауны края. Изучение ископаемых рыб без хорошего знакомства с остеологией совершенно немыслимо. Полезным пособием для ознакомления с морфологией чешуи может служить «Атлас чешуи пресноводных костистых рыб» Г. Г. Галкина (1958).
Уродливые рыбы, например с необычайно удлиненными или укороченными плавниками, с чрезвычайно малым или большим числом чешуи в боковой линии, с ненормальным отсутствием одного или нескольких плавников (чаще такая ненормальность встречается у брюшных плавников), или помеси, должны быть исследованы с особой тщательностью. Нужно сделать подробное описание, произвести многочисленные промеры по схеме того семейства или рода, к которому уродливая рыба относится. Цифры промеров таких рыб не следует заносить в таблицу наряду с другими обыкновенными рыбами, а вести отдельно от них.
В новых ихтиологических работах все большее внимание отводится анатомическим признакам. В упомянутых книгах А. Н. Световидова о трескообразных и сельдевых, В. В. Барсукова о зубатках и Д. Н. Талиева о бычках приложены целые атласы костей черепа и позвоночника этих рыб, что имеет значение для различия родов и видов, также даны рисунки костей висцерального скелета, жаберной крышки, поясов конечностей.
Анатомические признаки рыб успешно могут служить делу распознавания видов и более мелких таксономических групп рыб. К числу подобных исследований можно, для примера, отнести статью Ф. С. Замбриборщ (1951). Автор на основании анатомических особенностей глоточного аппарата и пищеварительного канала составил определительную таблицу для различения молоди трех. видов черноморских кефалей (Mugil saliens, М. auratus и М. cephalus).
Признаки помесей надо сравнивать с признаками исходных форм. Замечательные работы Н. И. Николюкина над гибридами карпа и карася (1938) показывают, что при скрещивании этих родов в некоторых признаках гибридов обнаруживается больше сходства с признаками материнскими (матроклиния), чем с отцовскими (патроклиния). Матроклиния обнаружена в числе позвонков и в некоторых других меристических признаках. Таким образом, встретив гибрида, у которого больше сходства с карпом, чем с карасем, мы можем допустить, что данный гибрид является результатом помеси самки карпа с самцом карася.
В природных условиях нередко встречаются межвидовые и межродовые гибриды рыб. Нельзя упускать ни одного такого случая без исследований и описаний. Но еще более существенно изучать гибридов искусственно выведенных, когда исходные родительские формы точно известны: это дает возможность проследить изменение и наследование признаков родителей и образование новых признаков.
Перед ихтиологами и рыбоводами стоит большая работа по гибридизации рыб на селекционно-генетической основе, так как это вызывается требованиями рыбного хозяйства, особенно на материковых водоемах, более всего на озерах и водохранилищах, наиболее доступных для управления качественным составом рыб. Часто бывает установлена и биологически обоснована необходимость изменения видового состава ихтиофауны. Это изменение может идти не только в направлении замены одного ‘вида другим, но и гибридизации рас и видов. В области гибридизации ихтиология имеет немаловажные достижения. Николюкин (1935, 1952) разработал методику межвидовой гибридизации, которой и следует пользоваться. Основательно разработана методика скрещивания красноперки с линем. Исследователь получил гибриды, которые по своим размерам превышали исходные формы-выявился гетерозис, что представляет несомненную хозяйственную ценность.
Н. И. Николюкин и Н. А. Тимофеева (1956) провели опыты по скрещиванию осетра и стерляди, белуги и стерляди на Волге и указывают на повышенную жизнестойкость гибридов. Можно ожидать, что промышленное выращивание таких гибридов представит практический интерес. На Урале промышленные результаты получены от гибридизации рипуса с чудским сигом (Нестеренко, 1957). Однако пока вопросам гибридизации рыб .уделяется недостаточное внимание.
Применение иммуносерологических реакций в ихтиологии
На протяжении многих лет иммунологические исследования концентрировались, главным образом, в области патологии инфекций. Подмеченная еще И. И. Мечниковым и его многочисленными учениками возможность использования иммунологических реакций для решения вопросов, не связанных с проблемой инфекции, долгое время не была надлежащим образом оценена.
С открытием в 20-х годах этого столетия групповых и типовых антигенов тканей иммунологические исследования стали широко применять в судебной медицине, санитарной экспертизе, переливании крови, а также для определения видовых и внутривидовых различий животных.
Необычайная чувствительность и точность этих методов исследования позволила использовать их в различных областях биологии: в систематике, эмбриологии, энтомологии, генетике, ихтиологии и т. д.
Благодаря усовершенствованию иммунологических методов исследования удалось обнаружить внутривидовые различия в самых различных группах животного мира: у грызунов (Хикс и Литли, 1931; Аврех и Калабухов, 1937; Ларина и Кулькова, 1959), птиц (Соколовская, 1936, 1938; Ирвин и Коль, 1936 а, б), дрозофил (Гершензон, Геронимус и Аврех, 1936), морских свинок (Линдштейнер, 1931) и т. д.
Иммуносерологические методы оценки внутривидовой изменчивости рыб привлекают за последние годы все возрастающее внимание и в зарубежных странах и в СССР; несомненно их значение будет увеличиваться по мере накопления опыта и дальнейшего совершенствования иммуносерологических методов. Эти методы позволяют установить различия не только между видами или подвидами, но и между внутривидовыми группировками. Это положение подтверждается исследованиями Нересгеймера (Neresheimer, 1908, 1909), который при помощи реакции преципитации доказал, что ручьевая форель весьма близка к лососю – тайменю и к озерной форме и, что не только морфологически, но и серологически эти формы представляют собой одну общую группу, тогда как американская радужная форель далека от европейской; Кодама (Kodama, 1913) при помощи реакции преципитации показал, что подклассы Chondrosteidae и Teleostei резко разграничены, а в последнем в свою очередь весьма резко разграничены семейства Clupeidae, Salmonidae и Cyprinidae; Джемерой (Gemeroy, 1943) провел на 31 виде рыб около 500 реакций; 0’Рурке (O’Rourke, 1959, 1960) проводил аналогичную работу при изучении родства близких видов и родов Gadus и Sebastes.
Основоположником изучения внутривидовой дифференциации у рыб является Д. Н. Талиев, обнаруживший серологические различия между расами байкальского омуля и хариуса. В ряде исследований (Талиев и Базикалова, 1934; Талиев, 1940) изложены результаты, полученные путем применения серологических методов для изучения истории байкальской фауны.
При изучении происхождения фауны озера Байкал Д. Н. Талиев и А. Я. Базикалова установили, что байкальская фауна смешанного происхождения, некоторые ее формы родственны северным морским рыбам, а часть их близко стоит к дериватам Каспийского бассейна.
В 1941 г. в работе «Серологический анализ рас байкальского омуля» Д. Н. Талиев использовал цельные и абсорбированные антисыворотки для реакции преципитации. В результате этих работ ему удалось установить три формы байкальского омуля. При определении родства сига с хариусом и ленком оказалось, что оба эти вида далеки от сига, но ленок более отдален, чем хариус.
Большой интерес представляет серологический анализ рас некоторых диких и одомашненных форм сазана (Талиев, 1946в). Оказалось, что степень отдаленности как чешуйчатого, так и разбросанного карпа по отношению к параванскому сазану одинакова, а родство амурских и параванских сазанов более отдалено, чем карпов и параванского сазана.
На основе своих многочисленных исследований Д. Н. Талиев пришел к выводу, что результаты серологических исследований не противоречат морфологическим данным, и иммуносерологические методы играют важную роль для определения начальных этапов внутривидовой изменчивости у рыб.
М. Г. Закс и М. М. Соколова (1961) использовали реакции кольцепреципитации и преципитации на агаровых пластинках по методу Оухтерлони для выяснения внутривидовых различий лососевых рыб Дальнего Востока. Вся работа была проведена на четырех стадах нерки, входящих на нерест в различные реки в разное время года. Результаты перекрестных реакций кольцепреципитации и преципитации на агаровых пластинках по методу Оухтерлони совпадали почти во всех случаях.
В результате проведенных исследований было установлено, что между локальными стадами нерки имеются определенные различия и родство, являющиеся доказательством наличия антигенно-серологической специфичности белков протоплазмы этих отдельных стад.
В 1961 г. Ю. П. Алтухов применил иммунологическую методику параллельно с цитофизиологической с целью определения влияния пола и возраста некоторых рыб на теплоустойчивость и степень серологической близости их мышечных белков.
По мнению автора, параллельное применение обеих методик, взаимно дополняя и контролируя друг друга, может сыграть большую роль при решении спорных вопросов систематики.
Для проведения серологического анализа Ю. П. Алтухов применил реакцию кольцепреципитации с сильно разведенными антисыворотками без истощения. Результаты серологического анализа оказались в полном соответствии с данными по теплоустойчивости. Пол и возраст различных рыб не оказывали существенного влияния на специфику их клеточных белков.
И. А. Балахнин (1961) использовал для определения внутривидовых различий крови форелей другую иммунологическую реакцию – гемоагглютинацию, основанную на выявлении внутривидовых антигенных различий в эритроцитах рыб. Ему удалось обнаружить внутривидовые различия не только у форелей, но и у карпа.
В 1963 г. Ю. П. Алтухов и В. С. Апекин при помощи реакции кольцепреципитации установили четкое серологическое различие между «крупной» и «мелкой» ставридами Черного моря, а также между этими формами и двумя видами Trachurus.
Различие между «крупной» и «мелкой» ставридой носит тот же характер, что и отличия этих форм от самостоятельных видов Trachurus. На основании этих данных можно считать, что формы ставрид, обитающие в Черном море, являются самостоятельными видами.
В последнее время появилось много американских, канадских, японских и французских работ, авторы которых пытаются устанавливать различия и родство между отдельными популяциями внутри одного вида по особенностям изо- или гетероагглютинации.
Известна работа Кушинга и Спрейг (Cushiing and Sprague, 1953), изучавших агглютинацию эритроцитов различных рыб посредством человеческой и других сывороток для установления родственных связей рыб по группоспецифическим свойствам их эритроцитов.
Риджвей, Кушинг и Дювалл (Ridgway, Gushing, Duvall, 1958) провели ряд работ с целью выявления различий популяции нерки серологическими методами.
Различия между отдельными популяциями нерки, добытой в разных районах, они устанавливали на основе неодинаковой степени агглютинации эритроцитов нерки свиной сывороткой.
В 1960-1961 гг. французский исследователь Ж. Ли (по Балахнину, 1962) при помощи реакций гемоагглютинации обнаружил различия между сардинами из трех районов одного и того же залива Средиземного моря.
Результаты серологического анализа согласовывались с данными по морфологии и биологии, и позволили утверждать, что сардины этого залива не однородны между собой.
Ряд исследователей выявляют внутривидовые группировки у рыб при помощи различных модификаций реакции агглютинации (Талиев, 1935; Кушинг и Дювалл, 1957; Сузуки, Мимото и Морио, 1959; Синдерман и Майрс, 1959; Риджвей и Клонтц, 1960).
Риджвей, Клонтц и Мотсумото (Ridgway, Clontz, Matsumoto, 1959) применили реакцию преципитации на агаровых пластинках по методу Оухтерлони для доказательства серологических различий между неркой Американского и неркой Азиатского побережий Тихого океана. При помощи этой реакции они обнаружили в сыворотке нерки Американского побережья 14 антигенов, а в сыворотке нерки Азиатского побережья-только 12. Таким образом, можно установить различие между нерками, выловленными в различных районах Тихого океана. Было обнаружено также присутствие особого антигена, характерного для половозрелых и созревающих самок нерки.
В течение последних лет методика постановки реакции преципитации в агаре получила широкое распространение при исследовании антигенного состава сыворотки животных и экстракта животных тканей.
Французский исследователь Кейванфар (Keyvanfar, 1962) применил реакцию преципитации в агаре по методу Оухтерлони при установлении различий между популяциями тунцов Атлантики и Средиземного моря. Он установил присутствие Одного антигена в сыворотке длинноперого тунца Атлантики, не содержащегося в сыворотке этого вида, обитающего в Средиземном море.
Автор приводит большое количество работ, которые делятся на две части: 1) исследование групп крови, 2) электрофоретическое и иммунологическое исследования сывороточных белков.
При изучении крови длинноперых тунцов Атлантики были установлены четыре группы крови. Эритроциты длинноперых тунцов из Средиземного моря также могут быть разделены на те же четыре группы. При этом была обнаружена четкая разница в распределении групп крови у длинноперых тунцов Атлантики и Средиземного моря.
Электрофоретический и иммуно-электрофоретический анализы дали возможность Кейванфару установить различия только между видами тунцов. Внутривидовые различия при помощи этого метода установлены не были.
Изучение белкового состава крови рыб, его изменчивости у разных экологических групп и при разном физиологическом .состоянии организма начато сравнительно недавно и развивается медленно. Нет данных о сезонных изменениях белкового состава крови проходных рыб и факторах, их определяющих. Материалы, относящиеся к рыбам пресных вод, показывают, что картина крови в течение зимы претерпевает весьма существенные изменения.
В последние годы некоторые зарубежные исследователи для целей систематики применяют метод электрофоретического разделения белков крови или других тканей на фильтровальной бумаге.
Так, Дриглон (Drihion, 1954) проводил электрофоретический анализ сывороточных белков крови самцов и самок угря и карпа.
При электрофоретическом исследовании белков плазмы крови у тресковых рыб К. М. Хайлов (1962) наблюдал отчетливые межродовые различия по количеству альбуминов и альфа-бетаглобулинов.
На основе полученных данных авторы пришли к выводу, что электрофоретические исследования имеют большое значение для выяснения степени родства или различия между отдельными таксономическими группами рыб.
Для изучения внутривидовой дифференциации осетровых рыб Каспийского моря Г. А. Амирханов использовал комплекс иммуносерологических реакций и метод электрофореза на бумаге.
В 1961 и 1962 г. были исследованы 116 экз. рыб (53 осетра “и 63 севрюги) ‘из Волги, Терека, Сулака, Самура и Куры в период весенней и осенней нерестовых миграций. От этих рыб были добыты антигены (сыворотки крови) и путем иммунизации кроликов были получены 44 иммунные сыворотки.
Для анализа полученного материала применили реакции кольцепреципитации с цельными и истощенными антисыворотками, преципитации на агаровых пластинках по методу Оухтерлони и реакцию связывания комплемента (РСК).
Методом электрофореза на бумаге было исследовано 28 сывороток крови севрюги и 25 сывороток крови осетра.
При исследовании были поставлены следующие задачи:
выяснить наличие антигенных различий сывороточных белков у локальных стад отдельных видов осетровых и изучить степень обособленности этих стад;
выяснить наличие антигенных различий сывороточных белков у биологических групп в пределах каждого локального стада и степень обособленности их между собой;
выяснить наличие или отсутствие внутривидовых различий! в составе белковых фракций сыворотки крови локальных стад и биологических групп у осетра и севрюги.
В результате проведенных работ удалось показать, что между отдельными локальными стадами имеются определенные различия, свидетельствующие о наличии антигенной разнокачественности их сывороточных белков. Эти особенности выявились как в реакции кольцепреципитации, так и в РСК, причем данные обеих реакций полностью совпадали. Антигенные различия у отдельных стад выявлялись не только при реакции с антигеном, которым пользовались для получения данной антисыворотки, но и с другими аналогичными антигенами от рыб того же стада.
Разумеется, реакции не во всех случаях выражены одинаково, что легко объяснить особенностями антисывороток, титры которых могут отличаться в зависимости от свойств продуцента. В отличие от кольцепреципитации и РСК реакция преципитации в агаре не дала ясных различий.
Иммуносерологический анализ различных стад осетров показал наибольшую степень родства между волжским и терским и между куринским и сулакским стадами. Самурское стадо занимает промежуточное положение, но ближе стоит к волжско-терской группе (рис. 19 и 20).
Наличие антигенных особенностей установлено и для локальных стад севрюги, входящих в те же реки (рис. 21 и 22).
При этом степень родства между локальными стадами у севрюг выражена в ином виде, чем у осетра. Куринское и терское стада проявили признаки большей близости, чем соответствующие стада осетров.
И для осетров, и для севрюг было лишь по одному случаю, когда один из антигенов проявил в обеих реакциях свойства, отличные от антигенов других рыб того же стада. Вероятно, такое явление связано с тем, что единичные экземпляры осетровых рыб заходят на нерест в необычную для них реку.
При иммуносерологическом сопоставлении биологических групп внутри одного и того же локального стада осетров и севрюг их антигенные свойства оказались совершенно тождественными (следует иметь в виду, что материал для анализов был взят лишь по I и IV биологическим группам).
По данным И. А. Баранниковой (1955), I биологическая группа по гистофизиологическим и биологическим признакам более близка к IV, чем ко II и III биологическим группам.
Осетровые антигены
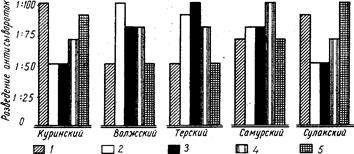
Рис. 19. Результаты реакции связывания комплемента между антисыворотками и антигенами различных локальных стад осетров. Осетровая антисыворотка:
1-куринская; 2-волжская; 3-терская; 4-самурская; 5 – сулакская.
Осетровые антисыворотки

Рис. 20. Результаты реакции кольцепреципитации между антисыворотками и антигенами различных локальных стад осетров. Осетровый антиген:
1 – куринский; 2 – волжский; 3 – терский; 4 – самурский; 5 – сулакский.
Очевидно, дифференциация I и IV биологических групп находится лишь в зачаточном состоянии, когда антигенные различия полков, затрагивающие наследственную основу более глубоко, пне не успели установиться.
Электрофоретический анализ сыворотки крови осетра и севрюги показал наличие четырех основных белковых фракций – альбуминов, альфа-, бета- и гаммаглобулинов.
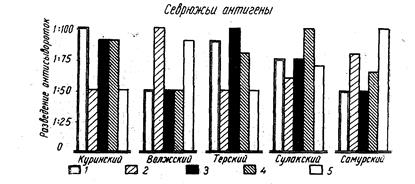
Рис. 21. Результаты реакции связывания комплемента между антисывороткой и антигеном различных локальных стад севрюг. Севрюжья антисыворотка:
1-куринская; 2- волжская; 3-терская; 4-сулакская; 5 – самурская.
Севрюжьи антисыворотки
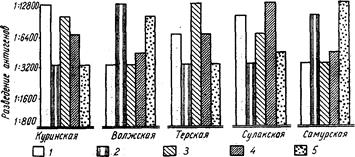
Рис. 22. Результаты реакции кольцепреципитации между антисыворотками и антигенами различных локальных стад севрюг. Севрюжий антиген:
1-куринский; 2- волжский; 3- терский; 4-сулакский; 5 – самурский.
Сравнительный анализ отдельных белковых фракций сыворотки крови осетра и севрюги внутривидовых локальных стад показал отсутствие существенных различий между последними, а между внутривидовыми биологическими группами в пределах одного стада наблюдались значительные сезонные различия.
При этом процентное содержание гаммаглобулинов у осенних рыб оказалось во всех случаях выше, чем у весенних. Очевидно, соотношение белковых фракций у осетровых-признак более лабильный, чем антигенные свойства, и может подвергаться изменениям в зависимости от- экологической ситуации. Антигенные свойства не обнаружили изменений в зависимости от возраста, пола рыбы, сезона и пр.
Вышеописанные опыты показывают, что комплекс иммуно-биохимических методов имеет очень большое значение в изучении характера внутривидовой дифференциации у рыб, а также в анализе сезонной динамики некоторых биологических показателей.
Следует отметить, что иммунитет рыб – один из наименее изученных разделов сравнительной иммунологии. Он представляет большой интерес не только при изучении родственных связей, но и для понимания эволюции механизмов защиты организма от заразных, болезней, а также для распознавания и предупреждения их.
Для диагностики краснухи карпов Г. Д. Гончаров (1949, 1950, 1951 а, 1959) применил одну из иммуносерологических реакций – реакцию агглютинации, в результате чего была подтверждена вирусная природа этой болезни. Помимо того, он (19516, 1962) проводил активную искусственную иммунизацию карповых рыб против краснухи.
При помощи реакции титрования комплемента В. И. Лукьяненко и Е. К. Миэсерова (1962) проводили сравнительно-иммунологическое изучение комплементарной функции крови четырех видов рыб. Им удалось обнаружить наличие резких различий комплементарной активности сыворотки не только у различных видов рыб, но и у отдельных представителей одного вида.
Однако необходимо отметить, что иммунологические методы в этой области пока еще применяются недостаточно.
Дальнейшее изучение этих вопросов поможет установить важнейшие закономерности сравнительной иммунобиологии, имеющей большое общебиологическое и эволюционное значение.
Проблемы иммунитета рыб в связи с перспективами развития рыбного хозяйства во внутренних водоемах имеют большое практическое значение.
Глава III
ПОЛ И ПОЛОВАЯ ЗРЕЛОСТЬ
Характеристика соотношения полов
Только у немногих видов рыб самцы и самки различаются по наружному виду (например, акулы и скаты). Некоторым рыбам свойственно половое различие во время нереста: нерестующие самцы лососей отличаются от самок более удлиненным рылом; у самцов линя брюшные плавники длиннее грудных;
у половозрелых самцов хариусов спинной плавник сильно увеличивается в высоту; у самцов полярных камбал чешуя обычно ктеноидная, у самок – циклоидная; есть и другие подобные примеры.
![]()
Рис. 23. Икорный щуп (по Беляеву, 1932).
В ихтиологических исследованиях определение пола рыб занимает большое место при работах по систематике и биологии рыб и при оценке состояния рыбных запасов. Эти исследования необходимы и для промысла. В .дальнейшем при более внимательном изучении внешних признаков самцов и самок рыб, несомненно, будет увеличено число видов рыб, пол которых различим по морфологическим признакам. Рыбаки различают самцов и самок судака по форме рыла. В нерестовый период самцы многих карповых рыб приобретают так называемый брачный «жемчужный» наряд чешуи (чешуя покрывается твердыми жемчужного цвета бугорками, наростами). Правда, такие бугорки бывают и на чешуе нерестующих самок, но у самок они менее обильны, чем у самцов.
Подобный нерестовый наряд можно встретить и у сигов. У рыб с развитыми половыми продуктами можно определять пол щупом (рис. 23), которым пользуются икорные мастера при установлении качества икры у осетровых рыб.
Обычно пол рыб определяют путем вскрытия. При наблюдениях на промысле (если рыбу потрошат) очень легко выяснить соотношение полов пойманных рыб в больших количествах. Наблюдатель может одновременно следить за несколькими резальщиками. Отметки самцов и самок делают сразу на раскрытом листке записной книжки: на левой стороне отмечают (черточками) самцов, на правой-самок. При таком способе в продолжение одного часа двое наблюдателей на амурском промысле успевали подсчитывать до 2000 рыб. Такого количества достаточно, чтобы вывести процентное соотношение полов данного вида в данный момент.
Соотношение полов у многих рыб близко 1 : 1 (по 50% того и другого пола). Но в разные биологические периоды это нормальное соотношение изменяется, и изменяется закономерно. У многих рыб в начале нерестового хода преобладают самцы, в конце хода – самки. Зная такую закономерность и следя за соотношением полов той или другой рыбы, можно делать прогнозы о повышении или ослаблении хода.
На промыслах, где заготавливают икру, такие прогнозы имеют большое значение (например, на дальневосточных лососевых промыслах).
Известно, что соотношение полов рыб зависит и от характера самого промысла. У многих рыб самцы (особенно в нерестовых стадах) мельче самок; если орудие лова крупноячейное, то оно отбирает крупных рыб, т. е. в основном самок, а зрелые самцы уходят из таких орудий. Поэтому для определения истинного соотношения полов рыб пробы нужно брать из таких орудий, которые в одинаковой степени улавливают как самцов, так и самок.
П. А. Дрягин (1952) формулирует следующим образом объем исследований по определению полового состава рыб:
1) определение полового состава молоди и половозрелых рыб;
2) половой состав отдельных генераций;
3) изменения полового состава в связи с возрастом;
4) половой состав в период нагула;
5) половой состав в период миграций;
6) половой состав в период нереста;
7) половой состав в период зимовки;
8) половой состав в уловах разными орудиями лова;
9) количество самок, которые могут быть использованы для сбора икры в рыбоводных целях и для заготовок товарной икры;
10) определение количества нерестовавших самок для расчета коэффициента возврата.
Обязательно следует фиксировать наименьшие и наибольшие размеры, вес и возраст половозрелых самцов и самок.
Стадии зрелости гонад и сравнительная оценка
отдельных шкал зрелости
Степень зрелости половых продуктов у отдельных видов рыб определяют различно. Существуют многочисленные схемы определения степени половой зрелости. Но однообразия в схемах даже по отношению к одному и тому же виду рыб пока нет. Вопрос этот освещен недостаточно, хотя уже много сделано особенно русскими исследователями: Вукотичем (1915), Киселевичем (1923 а и б), Филатовым и Дуплаковым (1926), Недошивиным (1928), Мейеном (1927, 1936. 1939, 1944), Кулаевым (1927, 1939), Трусовым (1947, 1949), Лапицким (1949). По этому же вопросу имеются ценные материалы в статьях многих наших ученых: Берга, Дрягина, Тихого, Вотинова, Наумова и др.
Единовременно нерестующие рыбы
Схема определения зрелости половых продуктов рыб, первоначально установленная старейшим ихтиологическим учреждением нашей страны – Астраханской ихтиологической лабораторией (ныне КаспНИРХ), описана К. А. Киселевичем (1923) в его «Инструкции для биологических наблюдений».
Схема определения зрелости гонад по Киселевичу
Стадия I. Неполовозрелые особи-juvenales. ( Латинские термины juvenis (множ. число-juvenes) и juvenalis (множ. число-juvenales) имеют различные значения: первый у римлян относился к организмам молодым, но уже возмужалым (по отношению к человеку-в возрасте свыше 20 лет); второй-к организмам юношеского возраста. Таким образом, при обозначении неполовозрелых рыб более правильно применять термин juvenales; отсюда – ювенальная (а не ювенильная) стадия.) Половые железы неразвиты, плотно прилегают к внутренней стороне стенок тела (по бокам и ниже плавательного пузыря) и представлены длинными узкими шнурами или лентами, по которым нельзя глазом определить пол.
Стадия II. Созревающие особи или развивающиеся половые продукты после икрометания. Половые железы начали развиваться. На шнурах образуются затемненные утолщения, в которых уже узнаются яичники и семенники. Икринки настолько мелки, что не видны невооруженным глазом. Яичники от семенников (молок) отличаются тем, что вдоль первых по стороне, обращенной к середине тела, проходит довольно толстый и сразу бросающийся в глаза кровеносный сосуд. На семенниках таких крупных сосудов нет. Половые железы малы и далеко не заполняют полости тела.
Стадия III. Особи, у которых половые железы хотя и далеки от зрелости, но сравнительно развиты. Яичники значительно увеличились в размерах заполняют от 1/з до 1/2 всей брюшной полости и наполнены мелкими непрозрачными, белесоватыми икринками, ясно различимыми невооруженным глазом. Если разрезать яичник и поскоблить концом ножниц по обнаженным икринкам, то они с трудом отрываются от внутренних перегородок органа к всегда образуют комки по несколько штук вместе.
Семенники имеют более расширенную переднюю часть и сужаются кзади. Поверхность их розоватая, а у некоторых рыб – красноватая от обилия мелких разветвляющихся кровеносных сосудов. При надавливании из семенников нельзя выделить жидких молок. При поперечном разрезе семенника края его не округляются и остаются острыми. В этой стадии рыба находится долго: многие виды (сазан, лещ, вобла и др.) – с осени до весны следующего года.
Стадия IV. Особи, у которых половые органы достигли почти максимального развития. Яичники очень велики и заполняют до 2/3 всей брюшной полости. Икринки крупны, прозрачны и при надавливании вытекают. При разрезе яичника и скоблении разреза ножницами икринки соскабливаются поодиночке. Семенники белого цвета и наполнены жидкими молоками, которые легко вытекают при надавливании брюшка. При поперечном разрезе семенника края его тотчас округляются, и разрез заливается жидким содержимым. Эта стадия у некоторых рыб непродолжительна и быстро переходит в следующую.
Стадия V. Текучие особи. Икра и молоки настолько зрелы, что свободно вытекают не каплями, а струёй при самом легком надавливании. Если держать рыбу в вертикальном положении за голову и потряхивать ее, то икра и молоки свободно вытекают.
Стадия VI. Отнерестовавшие особи. Половые продукты выметаны совершенно. Полость тела далеко не заполняется внутренними органами. Яичники и семенники очень малы, дряблы, воспалены, темно-красного цвета. 11ередко в яичнике остается небольшое количество мелких икринок, которые претерпевают жировое перерождение и рассасываются. Через несколько дней воспаление проходит, и половые железы переходят в стадию II-III.
Если половые продукты находятся на промежуточной стадии между какими-либо двумя из шести описанных стадий, или часть продуктов развита больше, часть меньше, или когда наблюдатель затрудняется точно обозначить стадию зрелости, то она обозначается двумя цифрами, соединенными знаком тире, но при этом та стадия, к которой ближе стоят по своему развитию половые продукты, ставится впереди. Например: III- IV; IV-III; VI-II и т. д. Основы этой схемы содержатся в схемах всех последующих авторов.
Шкала зрелости воблы и леща (по В. А. Мвйену и С. И. Кулаеву)
Самки воблы и леща (Эта шкала не применима для порционно нерестующего леща,)
Стадия I (ювенальная). Пол невооруженным глазом не различим. Половые железы имеют вид тонких прозрачно-стекловидных тяжей. На поверхности кровеносные сосуды или совсем отсутствуют или очень плохо различимы. В расщепленной железе при слабом увеличении под микроскопом видны отдельные яйцеклетки.
Эта стадия встречается у очень молодых особей в возрасте около I года.
Стадия II. Яичники имеют вид прозрачно-стекловидных тяжей желтовато-зеленоватого цвета. Вдоль яичника проходит тонкий кровеносный сосуд с очень мелкими ответвлениями. Икринки различаются невооруженным глазом или при помощи лупы. Они плотно прилегают друг к другу и имеют форму неправильного многогранника с округлыми углами. Процент веса яичников от веса всей рыбы составляет в среднем у воблы 0,77, у леща-1,21.
Стадия III. Яичник имеет округлую форму, немного расширяющуюся в головной части. На протяжении всего яичника невооруженным глазом видны икринки неодинаковой величины, многогранной формы (как в стадии II).
Кровеносные сосуды, расположенные вдоль яичника, хорошо развиты и имеют многочисленные ответвления. Процент веса яичника от веса всего тела рыбы в среднем составляет у воблы 3,26, у леща -4,1.
Стадия III у рыб встречается с конца августа до начала октября.
Стадия IV. Яичник сильно увеличился в объеме, и занимает большую часть брюшной полости. Икринки имеют неправильную многогранную округлую форму и тесно прилегают друг к другу.
При разрушении оболочки яичника икринки принимают шарообразную форму, так как не подвергаются больше давлению внутри яичника. Икринки плотно держатся в тканях яичника. В конце IV стадии (весной) в икринках видно невооруженным глазом ядро в виде небольшого пятнышка. Яичник обладает плотной оболочкой и отличается упругостью. Сильно развиты кровеносные сосуды с многочисленными ответвлениями. Стадия IV начинается с конца сентября или середины октября и продолжается до апреля-мая, т.е. до нереста. Процент веса яичника от веса тела всей рыбы осенью у воблы в среднем 8,3, весной – 20,9, у леща – 11,6.
Переход к стадии V (стадии полной зрелости) характеризуется появлением сначала отдельных прозрачных икринок, а затем небольших групп зрелых, прозрачных икринок. Потом зрелыми икринками заполняются целые участки яичника. Первое появление прозрачных икринок в яичнике показывает, что полная зрелость наступит в ближайшее время. Это состояние яичника обозначается как IV – V. Переходная стадия IV – V является кратковременной.
Стадия V. Яичник достигает полной зрелости и наполнен текучей икрой, выделяющейся при самом легком надавливании брюшка и даже при опускании рыбы хвостом вниз. Икринки прозрачные и имеют правильную шарообразную форму.
В самом начале стадии V икра хотя и прозрачна, но с трудом выделяется при надавливании. Затем наступает полная зрелость.
Стадия V у воблы и леща встречается в апреле – мае или начале июня.
Стадия VI. Яичник сильно уменьшился в размере и имеет дряблый вид, мягок на ощупь, багрово-красного цвета. Оболочка плотная крепкая. В яичнике встречаются редкие невыметанные икринки, часто беловатого цвета. Процент веса яичника от веса всего тела рыбы составляет в среднем у воблы и леща 1,3.
Переход стадии VI во II совершается постепенно в течение 1-1,5 месяца. По мере исчезновения пустых фолликулов и оставшихся после нереста икринок яичник постепенно из багрово-красного становится сперва розовым, затем розовато-стекловидным и, наконец, приобретает желтовато-зеленоватый цвет.
У повторно созревающих рыб после VI стадии наступает не II, а III стадия.
Самцы воблы и леща
Стадия I. То же, что и у самок.
Стадия II. Семенники представляют собой два тонких округлых тяжика почти такой же длины, как у зрелых семенников. Они мутные, бледно-розового или сероватого цвета. Кровеносные сосуды плохо заметны. Вес семенников очень мал и составляет у воблы в среднем 0,34% веса рыбы, а у леща 0,25%. Стадия II встречается в конце июля и в августе.
Стадия III. В начале стадии III (обычно сентябрь) семенники немного круглее, чем на предыдущей стадии, розовато-серые, упругие. В среднем они составляют 0,9% веса тела у воблы и 0,7% у леща.
В октябре-ноябре семенники матово-желтые, объем их значительно возрастает, а вес семенников у воблы составляет уже 2,25% и у леща 1,5% веса тела. В феврале – марте семенники достигают максимального размера, становятся упругими и приобретают розоватый или белый цвет. Молоки еще не выделяются при надавливании на брюшко. Даже при разрезании семенника они не выступают и не оставляют следов на бритве. Края разреза не сливаются и остаются заостренными. Вес железы достигает своего максимума и составляет в среднем 7% веса тела для воблы и 2,5% для леща. Эта стадия длится почти четыре месяца, затем цвет, объем, и вес железы резко меняются.
Стадия IV. Семенник вступает в период созревания. Макроскопические железа почти такая же, как на предыдущей стадии, но мелочно-белого цвета, и не так упруга. Проток еще не заполнен молоками. При надавливании на железу или на брюшко рыбы выступает густая капля молок. При разрезании семенника края разреза сплываются и выделяются густые молоки. Вес железы такой же или немного меньше, чем на предыдущей стадии.
Стадия IV встречается в апреле.
Стадия V. Семенники в состоянии полной зрелости и представляют собой два набухших эластичных мягких тела, равномерно мелочно-белого, чуть кремового цвета. На брюшной стороне тонкая прерывистая ниточка брюшного кровеносного сосуда. По мере выбрасывания спермы семенники становятся значительно тоньше, мягче, дряблее. Это изменение особенно резко сказывается в хвостовом отделе, где семенники буровато-розового цвета. В начале стадии очень резко выступает проток, который к концу стадии становится дряблым и розоватым, но остается хорошо различимым. В начале стадии молоки сами текут из невскрытой рыбы, в конце молоки еще вытекают при надавливании. Вес семенников в начале стадии у воблы около 7%, у леща 2,5%, в конце стадии – у воблы 3,4% и у леща 1 %.
Стадия V у воблы и леща встречается в апреле-июне.
Стадия VI (выбой). Семенники совершенно освободились от молок и представляют собою два тонких вялых тяжа. В поперечном сечении они угловатые, розового или буроватого цвета. Кровеносные сосуды видны плохо. Вес семенника резко снижается и составляет всего 0,5% веса тела у воблы и 0,4% у леща.
Стадия VI у леща встречается в июле.
Шкала зрелости окуня (по Мейену и Кулаеву, с сокращениями)
Самки окуня
Стадия I (ювенальная). Яичник одиночный и представляет собой небольшое прозрачное удлиненное тело, в котором нельзя различить невооруженным глазом отдельные икринки. Он бледно-желтого цвета с зеленоватым оттенком, стекловидно-прозрачен. По поверхности проходят небольшие кровеносные сосуды с мелкими разветвлениями.
Ювенальная стадия продолжается до середины второго лета жизни окуня.
Стадия II. Яичник стекловидно-прозрачен. Икринки очень мелкие, различимы невооруженным глазом, иногда при помощи лупы. Цвет бледно-желтый с зеленоватым оттенком.
Стадия II наступает у неполовозрелых особей в середине второго лета жизни и продолжается до середины следующего года. У половозрелых особей она наступает после окончания VI стадии и продолжается до августа, у таких особей ее следует обозначать как II-III.
Процент веса яичника от веса всего тела рыбы равняется в среднем 2,1%.
Стадия III. Яичник теряет прозрачность. Ясно видны отдельные круглые икринки, тесно включенные в ткани яичника. Цвет бледно-желтый. Стадия III у окуня начинается в августе и продолжается до октября. Процент веса яичника от веса всего тела рыбы равняется в среднем 3,5%.
Стадия IV. Яичник занимает большую часть брюшной полости. Икринки неправильной многогранной формы (при разрушении оболочки яичника становятся шарообразными), плотно связаны с тканями яичника. Цвет желтый. Стадия IV у окуня начинается с октября и продолжается до середины марта или начала апреля. Процент веса яичника от веса всей рыбы в октябре в среднем 8,8%, в феврале 13%, в марте-апреле-26,4%.
Стадия V. Икра текучая и выбрасывается в один прием. Стадия V наступает в конце марта или в апреле.
Стадия VI. Яичник сильно сжался вследствие спадения стенок. Мягок на ощупь. Красновато-серого цвета. Оболочка яичника сильно сжалась и утолщались. При разрезе яичника невооруженным глазом видны поперечные яйце несущие пластинки. В незначительном количестве встречаются невыметанные икринки. Процент веса яичника от веса всей рыбы-2,7%. Стадия VI у окуня продолжается в среднем в течение одного месяца после икрометания.
Самцы окуня
Стадия I (ювенальная). Половая железа в виде двух очень тоненьких и коротких стекловидных бледно-розовых полосок.
Стадия II. Семенники имеют вид двух тонких округлых тяжиков, мутного бледно-розового цвета. Длина их равна ‘/з развитого семенника. Вес семенника очень мал и в среднем составляет 0,2% общего веса тела рыбы.
Стадия II у рыб встречается в июне.
Стадия III. Семенники упругие розовато-серого цвета, увеличены в объеме и занимают половину полости тела. В начале стадии III (июль) вес их составляет 0,35% общего веса рыбы, а позже (к началу августа) -0,7% и, наконец, к концу стадии (к декабрю) – 2%.
К этому времени семенники почти достигают длины зрелой железы и имеют вид упругих, довольно толстых тяжей бледно-желтого и даже почти белого цвета. Молок еще нет. При разрезе края не сплываются и остаются заостренными. На бритве не остается мазка молок. Вес в среднем составляет 2% веса рыбы.
Стадия IV (созревание). Семенники очень крупные, почти достигают нормального размера зрелой железы и имеют молочно-белый цвет. Занимают всю полость тела. При разрезе на бритве остаются мазки молок, а иногда (на несколько более поздней стадии) при надавливании выступает густая капля молок. Вес семенника от 6 до 8% веса тела рыбы.
Стадия IV у окуня встречается с декабря до начала апреля.
Стадия V. Семенники в состоянии полной зрелости, достигают максимального размера, очень набухшие с гладкой, напряженной, эластичной поверхностью, мелочно-белого цвета. При надавливании на брюшко рыбы обильно выступают жидкие молоки. Вес железы достигает максимума и составляет 9% веса рыбы.
По мере выбрасывания молок семенники заметно спадаются, по объему равны приблизительно ‘/4 их объема в стадии зрелости; становятся дряблыми, морщинистыми, розового цвета, а в хвостовом отделе даже красного цвета. При надавливании еще выступают молоки. Вес железы резко падает и достигает в среднем 1,6% веса рыбы.
Стадия V у окуня встречается в апреле – мае.
Стадия VI (выбой). Семенники совершенно свободны от молок и представляют собой два тонких и вялых тяжа. Сильно укорачиваются и приближаются по величине и форме к стадии II, имеют буроватый цвет. Вес тоже приближается к стадии II и составляет в среднем 0,6% веса рыбы.
Стадия VI у самцов окуня встречается в конце мая.
Степень половой зрелости рыб прежде определяли невооруженным глазом. По существу это наиболее легкий, быстрый и практический способ, но он не дает ясной картины полного цикла развития гонад. Поэтому стали появляться работы, где степень зрелости яиц и спермы рыб описывается на основании микроскопического исследования. Микроскопический элемент внесен в схему Мейена для окуня, воблы, леща и др. Еще большее значение имеет гистологическое обоснование шкал для определения половой зрелости рыб. Подобная шкала предложена В. 3. Трусовым (1949а) для судака. В ней довольно подробно перечислены признаки, подмеченные невооруженным глазом, признаки, которые заметны под лупой, и гистологические.
И. И. Лапицкий (1949) предложил впервые шкалу зрелости половых продуктов для сигов, с учетом макроскопических и микроскопических признаков. Шкала Лапицкого написана очень ясно и вполне пригодна при полевых работах. Автор свою шкалу называет «промысловой».
Шкала зрелости половых, продуктов сига лудоги (по Лапицкому с сокращениями)
Стадия I (ювенальная). Яичники в виде двух валиков длиною 1- 1,5 см, продолжающихся нитевидными тяжами по бокам плавательного пузыря. Пол простым глазом не различим. Но под лупой или при малом увеличении микроскопа видны икринки. От семенника яичник отличается наличием крупного кровеносного сосуда и пластинчатым строением. Гонады бледно-розового цвета. Эта стадия продолжается до середины второго года жизни сига (1+).
Стадия II. Яичник в виде двух продолговатых тяжей длиной 3-5 см, округленных в головной части и сильно сужающихся в хвостовой. Цвет светло-розовый или слабо-оранжевый. Кровеносный сосуд, идущий вдоль железы, имеет многочисленные мелкие ответвления. Икринки различимы невооруженным глазом. Описанные признаки II стадии характерны для особей, еще не достигших половой зрелости, т.е. ни разу не участвовавших в нересте и длиться эта стадия до четвертого года жизни (3+). У самок, достигших половой зрелости и уже участвовавших в нересте, после выбоя икры наступает II стадия, которая макроскопически не отличима от описанной (гистологически отличима).
Стадия III. Яичники занимают от 0,50 до 0,75 длины полости тела. Икринки хорошо различимы простым глазом. Крупные икринки ярко-оранжевого, мелкие – светло-оранжевого или беловатого цвета. Яйценесущие пластинки яичника легко отделяются одна от другой и на каждой пластинке видны кровеносные сосуды.
Продолжительность стадии: с начала февраля до начала – середины октября.
Стадия IV. Яичники занимают всю полость тела. Икринки крупные, но среди крупных видны мелкие икринки.
Продолжительность стадии не более 15-20 дней (середина октября- первые числа ноября).
Стадия V. Период текучего состояния половых продуктов.
Стадия VI. Яичник в виде двух дряблых, сморщенных пластинок багрово-красного цвета. Много мелких икринок, изредка встречаются и крупным невыметанные икринки.
Продолжительность стадии: 1,5-2 месяца (первая половина ноября- декабрь).
Порционно нерестующие рыбы
Многими исследователями было подмечено, что у некоторых рыб период нереста занимает продолжительное время, и что у самок во время нереста размеры икринок различны. Но такие факты ранее принимались то за подход отдельных стад одного и того же вида, то более мелкие икринки вовсе не засчитывались в количество, подлежащее выметыванию в текущем году. Потом было установлено, что есть рыбы с единовременным икрометанием и рыбы с икрометанием порционным, другими словами, единовременно нерестующие и порционно нерестующие рыбы. По мнению П. А. Дрягина, есть виды рыб и с переходными признаками в характере нереста (Дрягин, 1949).
К. А. Киселевичем была установлена порционность икрометания для каспийских сельдей и в связи с этим предложена следующая схема определения стадий зрелости их гонад (Киселевич, 19236).
Стадии зрелости гонад у каспийских сельдей (по Киселевичу)
К. А. Киселевич указывает, что икрометание у каспийских сельдей происходит не сразу, а в три приема. Сначала выметывается одна порция икры, а остальная икра в стадии III, несозревшая, остается в яичнике и постепенно созревает в течение одной – полутора недели, проходя стадии IV и V. Когда и вторая порция выметана; то в яичнике остается последняя, третья порция в стадии III, которая дозревает в такой же срок и выметывается. Только после этого снова наступает полная стадия VI.
Для обозначения того, что первая порция икры уже выметана, впереди обозначения зрелости икры второй порции пишется в скобках римское VI; например: (VI) – IV означает, что первая порция икры выметана, а вторая находится в стадии IV. Если уже выметаны две первые порции, то в скобках ставятся две шестерки. Например, (VI-VI)-III, или (VI-VI)-V: первое означает, что рыбой выметаны две порции икры, а третья находится в стадии III; второе означает, что выметаны две порции, а третья в стадии текучести. Таким образом, весь период развития половых продуктов и икрометания у сельдей будет:
1) неполовозрелые (juvenales), стадия I;
2) первая порция икры, стадии: II, III, IV, V, VI-III;
3) вторая порция икры, стадии: (VI)-III, (VI)-IV, (VI)-V, (VI)-VI-III;
4) третья порция икры, стадии: (VI, VI)-III, (VI, VI)-IV, (VI, VI)’-V, (VI, VI) или просто VI, затем снова идет стадия III и т. д.
Распознавание первой, второй и третьей порций довольно затруднительно и удается только после некоторого навыка. Для облегчения даны следующие указания:
а) Первая порция икры всегда заполняет всю полость тела и распирает брюшко; семенники больших размеров и толсты. В стадии IV среди зрелых прозрачных икринок ясно видны мелкие, непрозрачные, более светлые незрелые икринки. Иногда среди икринок удается невооруженным глазом рассмотреть две группы-более крупные и более мелкие. Под лупой эти различия видны очень хорошо.
б) Вторая порция икры даже в период полной зрелости уже не заполняет всей полости тела, в которой замечается как будто пустота. Брюшко не распирается так сильно; яичники при той же длине, что и в первый раз, уже не так толсты и объемисты. В стадии IV среди зрелых икринок видны и мелкие, но их заметно меньше, и все они одинаковых размеров.
в) Третья порция еще меньше заполняет полость тела. Брюшко не раздувается, яичники длинны, но сравнительно тонки. В стадии IV и V среди зрелых икринок уже совершенно не замечается мелких, незрелых.
У самцов отдельные периоды икрометания выражены еще менее резко, и различить их гораздо труднее. Единственным указанием может служить степень опорожнения семенников:
а) в первый период весь семенник почти одинаковой ширины на всем своем протяжении;
б) во второй период задняя треть семенника уже опорожнена, передние же части еще широки и мясисты;
в) в третий период только передний конец семенника мясист и широк, задняя же часть опорожнена и имеет вид трубки.
Вообще у самцов общая картина зрелости семенников сильно маскируется тем, что при разной степени их зрелости можно выдавить хотя бы одну каплю молок. В период икрометания молок выдавливается больше, в промежуток между двумя порциями – меньше.
Полная шкала зрелости половых продуктов у сельдей, применявшаяся Волго-Каспийской рыбохозяйственной станцией, изложена в инструкции, опубликованной В. А. Мейеном (1939). Различаются шесть стадий зрелости: I-юношеская, II-стадия покоя (половые железы достигли нормальных для половозрелых особей размеров, но половые продукты еще не развиты), III- стадия развитых половых продуктов, IV-стадия созревания, V – половые продукты вполне созрели и VI – стадия выбоя.
П. А. Дрягиным (1939) разработана схема обозначения стадий половозрелости для карповых рыб, имеющих порционное икрометание. Схема П. А. Дрягина для уклей такова:
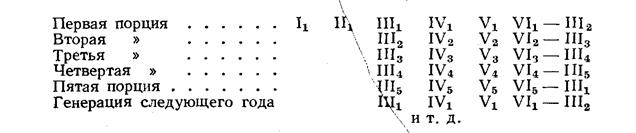
Римскими цифрами обозначены стадии зрелости икры по общепринятой шестибалльной схеме, арабскими указан порядок порций (выбоя).
В. А. Мейен (1940) рекомендует и макроскопическое исследование яичников по таким признакам: отношение веса половых желез к весу тела всей рыбы, степень прозрачности всего яичника и отдельных икринок, видимость икринок невооруженным’ глазом, видимость невооруженным глазом ядра в икринках, степень легкости выделения половых продуктов и общая форма половых желез. Кроме того, нужно указывать цвет половых желез, отличительные особенности их оболочки, упругость половых желез и степень развития кровеносных сосудов.
Очень много полезных сведений по половым циклам рыб дано в работах проф. П. А. Дрягина (1949, 1952 и др.).
Приведенные шкалы определения половой зрелости рыб (помимо этих шкал существуют и другие) не достаточно отчетливо характеризуют действительное состояние половых продуктов отдельных видов (или групп видов) рыб в отдельных промысловых районах. Не закончена морфологическая характеристика половых продуктов при разных степенях их развития (возрастающие размеры яиц, картина расположения кровеносных сосудов в гонадах при разных степенях развития половых продуктов, макроскопическая картина развития семенников и др.), а изучение гистологии яичников и семенников при отдельных стадиях развития только еще начато. Многие шкалы трудно применять в таких случаях, когда требуется быстро и на большом материале выяснить степень зрелости (например, для прогноза сроков подхода, рыб к нерестилищам и сроков самого нереста).
Поэтому необходимо разработать |более или менее универсальную шкалу, которой можно было/бы пользоваться и ихтиологу в полевых условиях и хозяйственнику. Такой шкалой, по моему мнению, следует пока признать ту, которая изложена проф. Г. В. Никольским (1944, 1963), и которая близка к первоначальной шкале Астраханской ихтиологической лаборатории.
I стадия. Молодые, неполовозрелое особи;
II стадия. Половые железы очень малого размера, икра простым глазом почти незаметна;
III стадия. Созревание, икра заметна простым глазом, наблюдается чрезвычайно быстрое увеличение веса половых желез, молоки из прозрачных становятся бледно-розовыми;
IV стадия. Зрелость, икра и молоки созревают (нельзя считать икру и молоки в этой стадии созревшими), половые железы достигают максимального веса, но при легком надавливании половые продукты еще не вытекают;
V стадия. Размножение, половые продукты вытекают уже при самом легком поглаживании брюшка, вес гонад от начала икрометания до его конца быстро уменьшается;
VI стадия. Выбой, половые продукты выметаны, и половое отверстие воспалено, половые железы в виде спавшихся мешков, обычно у самок с единичными оставшимися икринками, а у самцов с остатками спермы.
При наблюдении над половой зрелостью рыб всегда, надо указывать, какая из схем зрелости применялась наблюдателем.
О схемах стадий половой зрелости рыб подробные сведения приводятся П. А. Дрягиным в его статье-о полевых исследованиях размножения рыб (1952).
Напомним, что определение зрелости рыб нужно и для систематики, потому что некоторые морфологические признаки рыб изменяются в зависимости от зрелости яичников и семенников (размеры челюстей у лососей, наибольшая высота тела, величина плавников).
Коэффициенты и индексы зрелости
Вес гонад становится одним из обязательных условий выяснения степени зрелости половых продуктов, и в современных работах все чаще и чаще приводится (по предложению Г. В. Никольского, 1939) коэффициент зрелости, под которым понимается отношение веса гонад к весу рыбы, выражаемое в процентах. Определяют общий вес рыбы (т. е. с неудаленными половыми железами), затем вынимают половые железы, их взвешивают и определяют, какой % составляет вес гонад от веса всей рыбы. Такой коэффициент зрелости, конечно, не вполне отражает истинное состояние половых продуктов, но все же служит существенным пополнением схем зрелости.
Формула для вычисления коэффициента зрелости
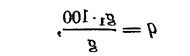
где q- искомый коэффициент зрелости;
g1 – вес гонад;
g –вес рыбы.
Коэффициент зрелости дозволяет следить за ходом созревания половых продуктов. Недостаток этого коэффициента состоит в том, что учитывается вес всей рыбы (вместе с кишечным трактом и с его содержимым). Этот вес колеблется в зависимости от наполнения кишечника, у рыб с наполненным желудком коэффициент зрелости будет преуменьшенным.
При наблюдениях за степенью зрелости половых продуктов у рыб с единовременным икрометанием П. А. Дрягин (1949) советует определять коэффициент зрелости, по меньшей мере, ежемесячно у особей половозрелого возраста и отдельно у особей, не достигших половой зрелости, причем должны быть отдельно учтены – максимальный показатель зрелости яичников перед самым началом нереста, показатель непосредственно после нереста и минимальный показатель перед началом нового периода созревания по окончании стадии VI.
У рыб с порционным икрометанием учитываются эти же показатели с ежемесячными наблюдениями и дополнительно следует обеспечить определение коэффициента зрелости перед первым, вторым и третьим выметом яиц, а также и непосредственно после вымета каждой отдельной порции.
Особое значение П. А. Дрягин придает максимальному коэффициенту зрелости, который характеризует период наибольшего развития гонад, что у рыб с единовременным икрометанием бывает незадолго до нереста (за одну-две недели), у рыб с порционным икрометанием – перед откладыванием первой порции икры. Хотя величина коэффициента – величина индивидуально колеблющаяся, все же она может характеризовать ход развития половых продуктов, свойственный отдельным видам рыб.
Определение максимального коэффициента зрелости яичников имеет значение теоретическое и практическое, например, для установления степени готовности яичников к нересту, для исчисления выхода икры в рыбоводных целях и при товарных заготовках, для учета плодовитости и сравнительной ее оценки у разных видов.
П. А. Дрягин предлагает также пользоваться коэффициентом зрелости для вычисления «индекса зрелости яичников». Под таким термином автор понимает «процентное соотношение коэффициента зрелости яичников, вычисленное в отдельные моменты их созревания и опустошения, к максимальному коэффициенту зрелости».
Пример. Коэффициент зрелости густеры в октябре-4,8. Максимальный коэффициент зрелости для этого вида определяется от 10,7 до 16,3, в среднем 13
![]() Индекс зрелости
Индекс зрелости
В сентябре индекс зрелости для густеры равен 29,0, а коэффициент зрелости – 4,0. Средние величины максимального коэффициента зрелости высчитаны еще для сравнительно небольшого числа видов рыб, да и установленные .средние максимальных коэффициентов еще могут быть уточнены. П. А. Дрягин относительно максимального коэффициента делает следующие предварительные выводы (сокращенно):
1. Каждому виду рыб свойствен свой показатель зрелости, более или менее отличный от показателя других видов.
2. Индивидуальная изменчивость коэффициента значительна-
3. Виды рыб с порционным икрометанием обычно имеют несколько меньший коэффициент зрелости.
При определении возраста рыб устанавливают и время наступления половой зрелости (способность к первому размножению). Исследованиями Фультона (Fulton, 1906) и особенно Дрягина (1934) доказано, что длина рыбы при достижении половой зрелости обычно в два раза меньше средней максимальной длины.
При определении возраста впервые нерестующих рыб необходимо помнить, что время наступления половой зрелости у одного и того же вида зависит от многих причин, и нужно его устанавливать для каждого исследуемого водоема. Наблюдения над созреванием яичников и семенников надо вести одновременно с метеорологическими и гидрологическими наблюдениями.
Общим правилом является более раннее половое созревание самцов. У лососей самцы становятся способными к размножению во время речного периода своей жизни, у самок этого не наблюдается. У морской камбалы (Pleuronectes platessa) Баренцева моря, как показали исследования Г. И. Милинского (1938), половая зрелость наступает у самцов преимущественно в возрасте 8-9 лет, а основная масса самок становится половозрелой не раньше 11-12-летнего возраста.
Сбор и фиксирование гонад
Сбор и фиксация половых желез производятся различными способами, но наиболее пригоден описанный В. А. Мейеном в его «Инструкции по определению пола и степени зрелости половых продуктов у рыб» (1939), откуда мы и заимствуем это описание (с изменениями).
От одной половинки половой железы-яичника или семенника берут три кусочка объемом около 0,5 см3 каждый, один кусочек отрезают от головной части железы, другой – от срединной и третий – от хвостовой, так как степень зрелости железы в названных участках может быть различной. Взятые пробы фиксируют сулемовым фиксатором или фиксатором Буэна.
Состав сулемового фиксатора: насыщенного водного раствора сулемы 100 см3 и ледяной уксусной кислоты 5-6 см3. Фиксация продолжается 3-4 ч, после чего объект переносят в 80-градусный спирт, в котором проба хранится. Перед исследованием пробу нужно поместить на одни сутки в слабый раствор йода на 96° спирту (цвет крепкого чая), чтобы не дать сулеме кристаллизоваться.
Состав жидкости Буэна: 15 частей насыщенного водного раствора пикриновой кислоты, 5 частей формалина 40-процентного и 1 часть ледяной уксусной кислоты. Фиксация продолжается 24 ч. После фиксации объект помещают на 1-3 ч в воду, которую несколько раз меняют, а затем кладут в 80-градусный спирт, где и хранят.
Есть и другие способы фиксации половых желез.
Хорошие результаты дает фиксатор из спирта и формалина: на 90 частей 70-градусного спирта берут 10 частей 40-процентного формалина, а также смесь Ценкера (5 г сулемы, 2,5 г двухромовокислого калия, 1 г сернокислого натра, 100 см3 дистиллированной воды). Перед употреблением прибавляют 5 см3 ледяной уксусной кислоты. Продолжительность фиксации до 24 ч.
Полезными руководствами при гистологических работах с икрой рыб могут служить книги Г. И. Роскина – Микроскопическая техника (1951) и Б. Ромейса под таким же названием (1953).
Заканчивая обзор методов определения степени зрелости половых продуктов рыб, необходимо сказать, что они еще не совершенны, потому что не дают детальной картины хода созревания яичников. Нужно продолжать работы по усовершенствованию методики определения зрелости гонад рыб.
Глава IV
ОПРЕДЕЛЕНИЕ ПЛОДОВИТОСТИ РЫБ
Значение изучения плодовитости рыб
Рыбы в сравнении с другими позвоночными животными поражают своей высокой плодовитостью, под которой разумеется количество яиц или икринок, откладываемых самкой в течение одного нерестового периода. Известны рыбы, которые за один нерест выметывают по несколько сот миллионов икринок (луна-рыба). Треска и угорь выбрасывают миллионы икринок, а рыб выметывающих сотни и десятки тысяч икринок, множество. Правда, есть и такие рыбы, плодовитость которых определяется небольшими количествами икры (до нескольких сот яиц – ручьевая форель, до нескольких десятков яиц – морская игла и до нескольких штук яиц – акула катран). Количество икры только до некоторой степени характеризует обеспеченность сохранения и умножения данного вида. Например, речной угорь выметывает до 10 млн. икринок, а горбуша-менее 2000 икринок, т. е. плодовитость угря в 5000 раз выше плодовитости горбуши. Оба эти вида только один раз в жизни оставляют после себя потомство, причем продолжительность жизни горбуши очень коротка, всего 1 ‘/2-2 года, а угорь живет около 10 лет. Между тем, едва ли поголовье речного угря в мировом масштабе в 5000 раз больше общего количества горбуши. Сельдь мало плодовитая рыба, а составляет основу мирового рыболовства. Луна-рыба, имеющая плодовитость до 300 млн. икринок, нигде не встречается стадами. Численность стада рыб обусловливается: степенью сохранности оплодотворенной икры, выживаемостью молоди, числом икрометаний, условиями размножения, условиями жизни взрослых рыб и т. п. Тем не менее количество откладываемой икры в сохранении вида – фактор весьма существенный.
Значение количества выметываемой рыбами икры необходимо для практических и научных целей.
Зная среднюю плодовитость разводимых рыб, рыбовод может составить реально осуществимый рыбоводный план завода или пункта и установить количество производителей, которое необходимо для искусственного оплодотворения.
Знание плодовитости необходимо и для суждения об эффективности естественного нереста рыб. При исследовании нереста и нерестилищ количество пришедших на нерестилища производителей и степень (количество) засева их икрой должны учитываться, так как это главнейшие факторы, влияющие на состояние запасов рыб. На определенную нерестовую площадь нужны определенное количество производителей и определенная густота размещения икры. У некоторых рыб (например, у дальневосточных лососей) можно учитывать количество идущих на нерест производителей и в зависимости от размеров и условий нерестовых площадей пропускать в реки нужное количество рыб. При этом необходимо знать плодовитость рыб.
Так как не всегда возможно провести наблюдения над самим нерестом, то для установления мест и сроков его, а также при работах, связанных с прогнозом численности стада рыб нужно проводить наблюдения над отложенной икрой. Даже количественную оценку размножения того или другого вида рыб для данного года можно давать по наблюдениям над выметанной икрой. Из советских ихтиологов методикой этих наблюдений над икрой много занимался проф. Т. С. Расе, инструкциями которого по сбору икры следует пользоваться (Расе, 1939 б).
Плодовитость рыб может служить отличительным признаком и при расовом изучении рыб. Осенняя кета, выделяемая в особую промысловую и систематическую группу как Oncorhynchus keta infraspecies autumnalis, отличается от летней большей плодовитостью.
При рыбоводных и селекционных работах необходимо учитывать разнокачественность икринок у разных особей и даже у одной и той же особи в разных частях яичника. Показатели разнокачественности икринок: диаметр (Дрягин, 1949), вес, окраска (Никаноров, 1959), размеры желтка.
Методика исследований, связанных с изучением плодовитости рыб, требует уточнения и разработки.
Различают индивидуальную плодовитость – общее количество икринок, выметываемых самкой за один нерестовый период; относительную – количество икры, приходящееся на единицу веса самки; рабочую – количество икры, идущее для целей искусственного оплодотворения (этот термин употребляется лишь в рыбоводстве); видовую и популяционную плодовитость.
Индивидуальная плодовитость
Для установления средней индивидуальной плодовитости необходимо располагать большим количеством цифрового материала и вести просчет икры надежным способом. Нужно брать икру в стадии наибольшего развития, но до момента наступления икрометания; икру нужно отбирать у самок различного возраста, и при просчете следует вести отдельный учет мелких недоразвившихся икринок, имея в виду, что такие икринки могут остаться невыметанными.
При взятии проб на плодовитость каждую самку нужно измерить и взвесить, а также взять чешую, плавниковый луч или другой объект для последующего определения возраста. Затем рыбу вскрывают, весь яичник взвешивают и отделяют пробу для просчета. Эта проба не должна быть большой: у лососей достаточно брать до 20 г, у других рыб – 5-10 г, у ряпушки – 0,5-2 г, т. е. чем мельче икринки, тем меньше навеска.
Пробу взвешивают на аптекарских роговых весах, кладут в баночку, снабжают этикеткой и заливают слабым (2-процентным) формалином (1 часть формалина на 19 частей воды). В соответствующем журнале записывают наименование рыбы, время и место поимки, орудие .нова, степень зрелости, длины тела: аb, ас, ad и оd, вес всей рыбы, икры и пробы. Надо оставить графы для вписывания количества икринок в навеске, во всем яичнике, диаметра икринок и для показателей возраста.
Для определения средних размеров икринок рекомендуется взять 10 икринок, расположить их по прямой линии, определить циркулем длину этой линии и разделив ее на 10, получить средний диаметр икринок. Еще лучше определять диаметр икринок с помощью окуляр-микрометра. Так как икра в воде набухает, надо измерять только что изъятые из яичников или фиксированные в формалине икринки.
Плодовитость рыб зависит от длины и веса рыбы.
В работе А. И. Ефимовой (1949) дана сводка плодовитости щуки обской, волжской и аральской (табл. 2).
Таблица 2
|
Длина, см |
Средняя индивидуальная плодовитость |
|||||
|
Обь |
Волга |
Аральское море |
||||
|
тыс. шт. |
% к общему количеству икры |
тыс. шт. |
% к общему количеству икры |
тыс. шт. |
% к общему количеству икры |
|
|
21-45 |
13 |
14,6 |
31 |
14,4 |
13 |
12,0 |
|
46-50 |
17 |
19,1 |
44 |
20,5 |
25 |
23,2 |
|
51-55 |
24 |
27,0 |
60 |
27,9 |
27 |
25,0 |
|
56-60 |
35 |
39,3<3 |
80 |
37,2 |
43 |
39,8 |
Приведенные данные я выразил по каждой группе в % к общей сумме икринок этих групп: обской, волжской и аральской щук, и получилось очень закономерное нарастание количества икры в зависимости от увеличения размеров тела рыбы.
Ф. Д. Великохатько (1941) на примере днепровского леща установил зависимость плодовитости от длины, веса и возраста рыбы. Семи- и восьмилетние лещи имеют наиболее высокую плодовитость. «У лещей более старших возрастов относительная плодовитость прогрессивно падает, тогда, как абсолютная плодовитость прогрессивно увеличивается». Самка 11 лет, весом 4315 г имела относительную плодовитость 136 икринок, а абсолютная плодовитость ее была 586530. У самки 6 лет, весом 413 г, относительная плодовитость-201,’ а абсолютная 332 850 икринок. Однако, у самых старых рыб наблюдается уменьшение количества икры.
При описании плодовитости все полученные материалы надо сводить в таблицы, для примера приводим табл. 3, взятую из работы М. И. Меньшикова (1936) по плодовитости сибирского осетра.
Таблица 3
|
Номер рыбы |
Вся длина рыбы, мм |
Возраст |
Вес рыбы, г |
Вес яичника . г |
Плодовитость ,шт. |
Относительная плодовитость |
Число яиц в1 г |
Отношение веса яичника к весу рыбы, % |
|
27 |
1422 |
20+ |
20400 |
3250 |
281222 |
14 |
89 |
15,9 |
|
26 |
1447 |
23+ |
16200 |
2060 |
174131 |
10 |
89 |
12,7 |
|
16 |
1460 |
24+ |
24000 |
3600 |
342612 |
14 |
103 |
15,0 |
|
13 |
1500 |
26+ |
24000 |
5800 |
419804 |
17 |
77 |
24,2 |
|
14 |
1500 |
26+ |
22500 |
3700 |
268546 |
12 |
83 |
16,4 |
|
21 |
1500 |
23+ |
24800 |
3540 |
349 752 |
14 |
131 |
14,2 |
|
46 |
1504 |
26+ |
31000 |
4300 |
217924 |
7 |
60 |
13,8 |
|
28 |
1509 |
24+ |
26000 |
4250 |
345653 |
13 |
89 |
16.4 |
|
Средняя |
1480 |
24 |
23613 |
3813 |
299 936 |
13 |
90 |
16 |
Примечание. Автор не приводит средних, но их давать необходимо, особенно когда число самок, от которых взята икра, превышает 10 шт.
Различные авторы составляют сводки по плодовитости рыб по-разному. Довольно наглядные таблицы но индивидуальной плодовитости приводит М. А. Летичевский (1946) для 6 промысловых рыб Аральского моря.
В табл. 4 мы приводим плодовитость щуки по М. А. Летичевскому.
Количество, икры он определял по навеске в 1 г.
Эта табличка может служить образцом при работах, когда нужно вскрыть зависимость количества икринок от размера, веса и возраста исследуемого вида рыб. Недостает в табличке лишь показателей числа исследованных самок каждой категории.
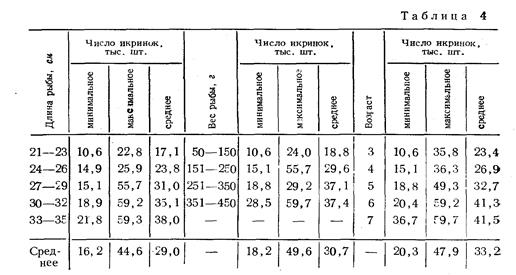
Так как у многих рыб половые железы развиваются неодинаково, например у мойвы в одном (левом) яичнике было насчитано 7860 икринок и другом (правом) – только 475 икринок (Суворов, 1931), и размеры икринок не всегда одинаковы, то навеску для просчета икры следует брать из разных мест и из обеих половинок железы.
Иногда применяется объемный метод определения плодовитости. В настоящее время этот метод допускается только применительно к просчету крупной икры, и все же точности здесь не достигается. В рыбоводстве объемным способом пользуются при счете икры лососевых и осетровых рыб. Взвешивают всю икру, затем берут 2 или 3 пробы и наполняют ими 25 см3 градуированной мензурки, точно просчитывают число икринок в этом объеме сосуда и по этой пробе определяют количество всей взятой из рыбы икры, объем которой известен.
Объемный способ просчета икры более подробно описан (по Францу, 1909), Е. К. Суворовым (1948) и состоит он в следующем.
Отделенные от оболочки икринки в 70%-ном спирту помещают в калибрированный сосуд, закрытый пробкой. Через отверстие в пробке вводят планктонную пипетку Гензена: жидкость старательно встряхивают для равномерного распределения в ней икринок, и в то же время в пипетку втягивается 0,5- 1 см3 спирта с икринками. Затем пробу распределяют с помощью кисточки на черной навощенной дощечке (10 см2), разграфленной на квадратные сантиметры. Полученные путем подсчета цифры записывают на разграфленную бумагу. Такие подсчеты нужно несколько раз повторить и исходить из средней.
Отсюда нетрудно рассчитать общее количество икринок. Например, морская камбала 8 лет, 51 см длины; икринки заключаются в 440 см3 спирта; в пробе (1 см3) подсчитано 776 икринок, следовательно, всего икры 776Х440=341 440 шт.
В работе П. А. Дрягина (1952) описан графический способ определения плодовитости по диаметру икринок и их общему объему. Измеряют диаметр у 50-100 и более икринок и вычисляют их средний диаметр. Определяют общий объем яиц, содержащихся в яичниках. Затем по графику Байера находят количество яиц в данном объеме.
Этот графический метод с успехом может быть заменен вычислениями по уравнению, предложенному В. А. Амосовым (Дрягин, 1952):

Описанным методом особенно удобно пользоваться в рыбоводной практике, когда нужно быстро определить количество икры.
Относительная плодовитость
Для определения относительной плодовитости берут общий иге рыбы (кг или г), определяют число икринок во всем яичнике и делят на вес рыбы. Сравнение относительной плодовитости допустимо лишь для отдельных стад одного и того же вида.
Относительная плодовитость важна в рыбоводстве. Имея расчет количества икры на 1 кг веса тела рыбы, можно, но очень грубо определить по весу самки и количество имеющейся в ней икры (количество икры зависит и от размера и возраста рыбы).
Рабочая плодовитость
Рабочая плодовитость рыб принимается рыбоводами, и введен этот термин после того, как рыбоводы установили, что они не могут получить такое количество способной к оплодотворению икры, которое соответствует индивидуальной плодовитости каждой самки. Полученная искусственным путем (путем отцеживания) икра тоже не вся остается живой и годной к оплодотворению. В руках более опытного рыбовода рабочая плодовитость будет выше, чем плодовитость той же рыбы, но взятой неопытным лицом. Никакой методики для установления рабочей плодовитости рыб пока не разработано, и мало случаев, когда приводят показатели рабочей плодовитости. У П. А. Дрягина (1948) есть упоминание, что рабочая плодовитость пеляди составляет около 73% от абсолютной плодовитости.
Плодовитость порционно нерестующих рыб
Порционное икрометание, когда рыба в течение одного нерестового сезона откладывает икру несколько раз, т. е. порциями, было подмечено у сельдей К. А. Киселевичем (1923, 1937), у карповых рыб П. А. Дрягиным (1939, 1952). Статьи П. А. Дрягина остаются основными руководствами при изучении порционного икрометания рыб.
Диаметр икры таких рыб имеет различные величины. У густеры перед моментом откладывания первой порции икры можно видеть четыре группы яиц (в мм) (Дрягин, 1939):

Среди икры, которая должна быть выметана в текущем году, есть немалое количество икринок с меньшими диаметрами – до 0,1-0,2 мм.
Четвертая (последняя или резервная) группа яиц, по мнению П. А. Дрягина, будет выметана в следующем году, и количество икринок, содержащихся в этой группе, нельзя засчитывать в плодовитость рыбы.
«При определении плодовитости порционно нерестующих рыб приходится различать плодовитость по каждому отдельному вымету, плодовитость по остаточной икре и общую, индивидуальную плодовитость. Последняя может быть определена до начала первого нереста по сумме всех икринок, включая крупные и все мелкие икринки; у особей последующих нерестов, т. е. второго, третьего и т.д. выметов, можно определить лишь остаточную плодовитость по учету крупной и мелкой остаточной икры» (Дрягин, 1939). .
Порядок составления ведомости по просчету икры у порционно нерестующих рыб (по Дрягнну) дается в табл. 5 .
Следует иметь в виду, что у некоторых мелких и скороспелых видов рыб, обладающих особым типом «неперемежающегося» овогенеза, фактически нельзя определить общую плодовитость без экспериментальных данных. У таких видев все время в течение нерестового периода идет пополнение созревающих овоцитов разных стадий развития гамет из так называемых резервных овоцитов. Например, П. К. Гудимович (1952) в период наблюдений над нерестом черноморской султанки с 4 по 20 июня 1951 г. отмечал у нее лишь три порции икры, тогда как Л. С. Овен (1961 а, 1961 б), наблюдая нерест черноморской султанки в аквариумах Карадагской биологической станции АН УССР, установила у этого вида ежесуточный нерест, повторявшийся до 60 раз за нерестовый сезон (при подкармливании).
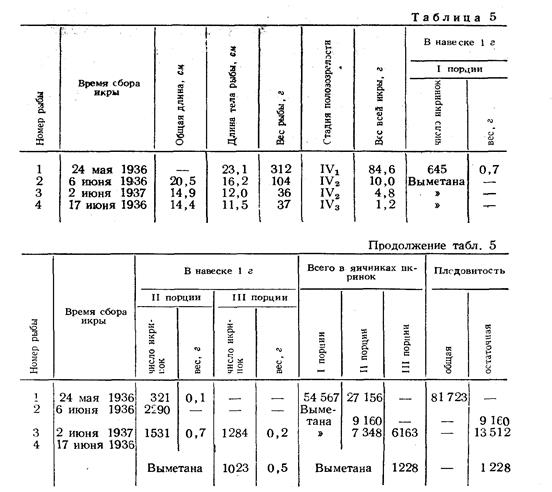
Видовая плодовитость
Видовая абсолютная плодовитость – сумма икринок, откладываемых самкой в продолжение всей жизни, видовая относительная плодовитость показывает суммарную производительность яиц на единицу веса тела рыбы за всю ее жизнь (Иоганзен, 1955).
В понятие о видовой плодовитости рыб значительную ясность внес С. А. Северцов (1941), который указал, что плодовитость, основанная только на просчете икринок без учета значения других констант, является плодовитостью кажущейся, и предложил особую формулу для определения относительного показателя видовой плодовитости.. Северцов считает, что плодовитость каждой популяции вида определяется наследственными константами размножения, обозначенными буквами:
r- среднее число детенышей на пару в год (годичный цикл j жизни животных);
j–средняя величина возраста первого плодоношения самки;
p– период между двумя последовательными деторождениями этой самки;
s-отношение числа самок к числу самцов.
Формула Северцова имеет вид
![]()
В применении к, рыбам г – индивидуальная плодовитость, j – возраст при наступлении половой зрелости, р – период между двумя икрометаниями.
Пользуясь такой формулой, А. И. Ефимова (1949) определила видовую плодовитость щук для двух разных районов: для Оби и для Волги (табл. 6). (В таблице А. И. Ефимовой не отмечено, какой принят показатель для соотношения полов.)
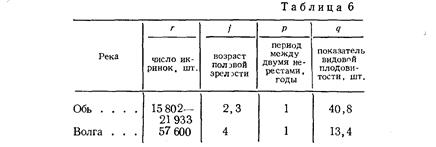
Приведенный расчет показывает, что у волжской щуки видовая плодовитость ниже, чем у обской, хотя индивидуальная плодовитость у волжской выше. Таким образом, повышение видовой плодовитости может идти или через повышение плодовитости индивидуальной, или через более короткие сроки полового созревания, или путем многократности размножения П. А. Дрягиным (1952) приводятся следующие показатели видовой плодовитости (в шт.) для ряда рыб бассейна Оби (вычисления В. А. Амосова):

Наименьшие показатели, пишет П. А. Дрягин, имеют виды. подвергающиеся слабой межвидовой конкуренции и ничтожному воздействию хищников (осетр, стерлядь), и хищные рыбы (нельма, щука). Наибольшие показатели свойственны рыбам, испытывающим острую межвидовую конкуренцию, пресс хищников, или подверженных большой естественной смертности (тугун).
Б. Г. Иоганзен (1955) указал, что в формуле определения показателя видовой плодовитости С. А. Северцова не учитывается продолжительность жизни особи, а в связи с этим и число нерестов. Поэтому им предложена следующая формула для определения относительного показателя видовой плодовитости:
![]()
где х – число икрометаний в течение жизни. Этим автором из формулы исключается показатель s.
Популяционная плодовитость
В. С. Ивлевым (1953) обоснована необходимость более точной оценки видовой плодовитости рыб с учетом локально выраженной возрастной неоднородности их стад. Им предложен показатель популяционной плодовитости (R), который вычисляется по следующей формуле (Дано в упрощенном виде.)
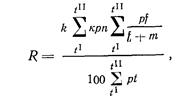
где t- возраст в годах;
tI возраст, при котором наступает половозрелость;
tII –возраст, при котором особи прекращают нереститься (максимальный возраст рыб в популяции);
р – относительная величина данной возрастной группы» выраженная в % от общего количества половозрелых особей;
п – абсолютная плодовитость одной самки данного возраста;
f– число самок в средней пробе;
m– число самцов в средней пробе;
k – число икрометаний в течение года.
Ю. И. Никаноров (1959) исчислил популяционную плодовитость по формуле В. С; Ивлева у ряда локальных стад ряпушки, обитающей в озерах Латвийской ССР и Получил следующие результаты:
|
Озеро |
Год |
Индивидуальная плодовитость, шт. |
Популяционная плодовитость, шт. |
|
Свента |
1953 |
4030 |
886 |
|
Свента |
1956 |
4360 |
655 |
|
Керза |
1957 |
12190 |
1194 |
|
Сивер |
1957 |
6630 |
843 |
|
Дризда . |
1956 |
11700 |
945 |
|
Разнаc |
1956 |
10700 |
1264 |
Он считает, что установленные различия в плодовитости можно рассматривать как одну из форм проявления внутривидовой биологической неоднородности ряпушки в зависимости от разных условий обитания и что чем больший процент в нерестовом стаде составляют младшие возрастные группы, тем выше популяционная плодовитость.
Г. В. Никольский и Т. Н. Белянина (по Никольскому, 1963) при подсчете плодовитости одного и того же стада рыб в разные годы считают возможным упростить формулу В..С. Ивлева, определяя плодовитость популяции рыбы на 1000 шт. самок при данном возрастном составе и данной средней плодовитости рыб каждой возрастной группы. Г. В. Никольский указывает, что этот показатель позволяет правильно оценивать плодовитость стада и его динамику для рыб с единовременным икрометанием.
Приступающие к сбору материалов по плодовитости рыб обычно задают вопрос: какое количество проб необходимо брать? Едва ли можно на такой вопрос дать ответ, пригодный для любого вида рыб. Количество икры зависит и от длины рыбы, и от веса, и от возраста; размеры икринок у рыб различны; у разных рыб навески икры для просчета нужно брать разные; одни исследователи ограничиваются определением плодовитости лишь нескольких штук самок, другие исследуют сотни рыб.
Необходимо определять плодовитость у каждой возрастной группы.
Возрастных промысловых групп обычно немного – менее 10. Нередко в промысловом стаде преобладают только 2-3 возрастные группы. Единичные особи самых старших групп не могут влиять на среднюю плодовитость вида, определение плодовитости таких отдельных самок имеет лишь частный интерес, как показатель максимальных количеств икры.
Исходя из таких соображений, можно рекомендовать брать 11 каждой возрастной группе по 10 яичников, т. е. всего для каждого вида соберется 100 проб (в том случае, когда возрастных групп 10, в действительности этих групп бывает меньше).
Глава V
НЕРЕСТ И НЕРЕСТИЛИЩА
Общие указания о наблюдениях над нерестом рыб
При полевых работах наблюдательный ихтиолог приобретает много сведений по поведению рыб в их нерестовый период, но часто эти сведения ихтиологами записываются настолько кратко, что по ним можно лишь установить сроки икрометания рыб, а использовать их для освещения многочисленных, весьма сложных биологических явлений, предваряющих и сопутствующих нересту рыб, нельзя. Нужно проследить и подробно описать поведение рыб перед началом нереста (соотношение полов, предбрачные изменения морфологических признаков, преднерестовое питание, проявление забот о подыскании нерестилищ и т. п.). Еще необходимее получить материалы по поведению рыб во время самого процесса икрометания (соотношение полов, брачные изменения признаков, брачная игра, процесс откладывания и сохранения рыбами икры и т. п.). Наконец, нужно собрать материал, относящийся к посленерестовому периоду рыб (скат рыбы, питание, общее состояние отнерестившейся рыбы).
Наблюдений над нерестом рыб в естественных условиях слишком мало, наиболее полно исследован нерест дальневосточных лососей. Стеллер, талантливый сподвижник Беринга, двести лет тому назад дал прекраснейшие сведения о нересте дальневосточных лососей по своим непосредственным наблюдениям на Камчатке. Затем И. И. Кузнецов и его сотрудники вновь и более тщательно проследили нерест той же группы рыб.
Е. М. Крохин и Ф. В. Крогиус (1937) провели обстоятельное исследование нерестилищ дальневосточных лососей на Камчатке. Они характеризуют гидрологические условия нереста и нерестилищ. Например, горбуша нерестует при скоростях течения от 0,3 до 0,6 м/сек при рН не ниже 7,0.
К. Ф. Телегин (1932) дал хорошие сведения об условиях преднерестовых и посленерестовых миграций беломорской наваги.
Советские ихтиологи обогатили науку работами по размножению рыб. Отличительная черта этих работ выражается в их экологической направленности. В работе проф. П. А. Дрягина (1949) приведена сводка данных о ходе нереста рыб на примере озера Ильмень. Этими данными как методическим руководством можно пользоваться при наблюдении нереста рыб и на других водоемах.
В работах В. И. Владимирова (1955) и В. И. Владимирова, II. Г. Сухойвана и К. С. Бугая (1963) даны полезные сведения о размножении рыб в условиях зарегулированного стока Днепра и, в частности, о нересте и нерестилищах рыб.
Чтобы изучить и понять биологию нереста рыб, нужно проводить тщательные и полные наблюдения в искусственных, возможно близких к естественным, условиях, например в садках и и аквариумах. Существует много конструкций живорыбных садков. Впервые мысль об устройстве садков для выдерживания производителей у русских ихтиологов осуществлена в 1909 г. (Кузнецов, 1912). Отказавшись от применения живорыбных садков, И. И. Кузнецов построил на одном из протоков р. Амура садок с естественным грунтом. Перпендикулярно к берегу в два ряда (ширина между рядами около 5 м) на протяжении 20 м через каждые 1-1,5 ж были забиты колья толщиной от 10 до 15 см. С внутренней стороны этих кольев была протянута на небольших шестах крупноячейная сетка из цинковой проволоки. Исследователю удалось выдержать производителей кеты до полного созревания, проследить процесс нереста – выметывание и оплодотворение икры, зарывание икры в грунт и т. п.
Есть немало и других вопросов, относящихся к нересту рыб. Накопился значительный материал, указывающий на не ежегодное икрометание некоторых рыб. Осенью, в период нереста, можно встретить взрослых сигов, которые имеют совершенно не развитые половые продукты. О подобном же явлении писал и С. В. Аверинцев (1933): «изучение нельмы и сиговых Лены дало мне определенные указания на то, что нерест, по крайней мере у некоторых видов, происходит вовсе не ежегодно».
Не ежегодно нерестуют и те лососи (например, озерные лососи), которые остаются живыми после первого нереста и будут готовиться к следующему нерестовому периоду. Ладожский озерный лосось после нереста настолько изнуряется и худеет. что нельзя допустить, чтобы он истомленным спустившись из реки в озеро весной, мог осенью того же года снова идти на нерест (Правдин, 1948). Однако решение вопроса о не ежегодном нересте в ряде случаев осложняется тем, что половозрелость не у всех рыб наступает в одном и том же возрасте, и потому в стаде рыб часто можно обнаружить особей одинакового возраста и размера с различной степенью половозрелости (is половозрелом и неполовозрелом состоянии).
Насколько свойственна рыбам гибель после нереста? Вопрос, который также имеет большое значение. Известно, что тихоокеанские лососи погибают после первого же нереста. Но высказываются мнения, что отдельные особи не гибнут. А. Г. Кагановский (1949) детально обсуждает вопрос о возрасте дальневосточной горбуши. До этой работы было одно мнение, что горбуша идет в реки на нерест на втором году своей жизни. А. Г. Кагановский приходит к заключению, что в некоторые годы с неблагоприятными для горбуши факторами часть ее стада может настолько отстать в своем развитии, что достигнет полового созревания только на третьем году жизни. Материалы А. Г. Кагановского, конечно, не могут быть использованы в качестве доказательства о выживаемости производителей горбуши после нереста, но они подтверждают, что горбуша половой зрелости может достигать не только на втором году жизни.
Широко распространенное мнение о массовой гибели волжской сельди (Caspialosa kessleri volgensis) после нереста теперь должно измениться, так как Н. П. Танасийчук (1948) привел неопровержимые данные о неоднократном икрометании этой сельди – она может нерестовать до четырех раз в жизни.
При исследовании нереста необходимо вести наблюдения не только над поведением нерестующих рыб, но и над температурой воды, состоянием уровня воды в водоеме, быстротой течения и над другими гидрологическими факторами.
Помимо дневника наблюдений, куда вписывают ежедневные данные по всем работам наблюдателя, в том числе и по нересту рыб, рекомендуется вести отдельный журнал или ведомость наблюдений над нерестом. Журнал ведется для каждого вида рыб отдельно. Размеры и форма журналов по нересту рыб могут быть различны, но для каждого вида рыб нужно указывать: пол и состояние половых продуктов, время поимки (месяц, день и час), длину и вес рыбы, температуру воды (в поверхностном слое и обязательно в зоне поимки рыбы), глубину водоема до дна на месте поимки и в зоне поимки. Журнал имеет примерно такой вид:
Наименование рыбы____________________________
Наименование водоема__________________________
Наблюдатель___________________________________
Время наблюдения: число и месяц начала наблюдений 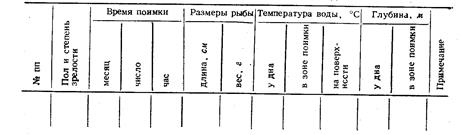
При обработке журнала по нересту берут и другие гидрометеорологические сведения. Желательно, чтобы в нерестовый журнал с самого начала нереста или еще до его наступления, т. е. с того времени, когда стадия половых продуктов рыбы приблизилась к полному созреванию, ежедневно вписывались нерестовые анализы (50 экз. самцов и самок, с разбивкой этого количества по трем основным группам нерестующих рыб: мелкие, впервые нерестующие, средние и крупные, старые). У каждой рыбы надо брать чешую или кости для определения возраста. Составление такого журнала за весь нерестовый период по отдельным видам рыб окажет громадную пользу при последующей характеристике нереста. Сроки и продолжительность нереста рыб определяются не только индивидуальными и видовыми свойствами рыб, не только гидрологическими факторами, но и общеклиматическими даже микроклиматическими.
Общеклиматические сведения можно получить в специальных географических и климатологических книгах и статьях. Кроме того, в нашей стране существует сеть метеорологических станций и водомерных постов, где ихтиолог может получить сведения по температуре воздуха и воды и другие метеорологические данные за ряд лет, а также указатели по колебаниям уровня воды. Эти сведения для ихтиолога необходимы. Но при изучении нереста рыб, и вообще биологии рыб, крайне важно знать не только такие общие сведения в виде средних годовых и месячных по району или области, но и получить такие же сведения по данному водоему или даже по участку водоема, где проходят ихтиологические работы. Поэтому ихтиологу приходится организовать гидрометеорологические наблюдения в пункте своих работ. Свои наблюдения надо сопоставить с данными соседней метеорологической станции.
Установление сроков и продолжительности нереста
Нерест у разных рыб протекает по-разному: у одних очень быстро – в 2-3 дня, у других – до месяца и более. Рыбы нерестуют весной (щуки и большинство карповых), летом (карась, некоторые осетровые и др.), осенью (большинство лососевых рыб), зимой (налим, треска, навага). Наблюдатель должен проследить ход нереста каждого промыслового вида рыб (отмечая его начало, разгар и конец).При наблюдении над сроками нереста, отдавая должное данным, опубликованным другими исследователями, все же необходимо продолжать сбор материалов по времени нереста. Есть указания, противоречащие общепринятым данным по этому вопросу. Например, всеми считается, что сиги откладывают икру осенью или в первой части зимы, о муксуне (Coregonus muksun) всюду сообщается, что он нерестует в октябре – ноябре. Но в. работе Е. В. Бурмакина (1938) об ихтиофауне Гыданского залива указывается, что в Гыданском заливе были обнаружены особи муксуна с текучими половыми продуктами в период с 25 апреля по 3 мая ( 1936 г.). Такое указание (которому не верить нет оснований) резко расходится с тем, что было до того времени известно о времени нереста муксуна.
В Швеции есть ряпушка с весенним нерестом.
Эти примеры показывают, что по нересту даже широко распространенных рыб исчерпывающих данных еще нет, и что нужны всюду более полные сведения, так как нерест рыб представляет собой не только теоретический интерес, но и промысловое значение.
Наблюдатель должен помнить, что в зависимости от состояния гидрометеорологических условий не только изменяются сроки нереста, но изменяется и самый нерест. Это особенно хорошо известно в отношении нереста карпа. В Ленинградской области нерест карпа отмечается от 19-20 мая до 13-14 июня (Савельев, 1941). В особенно холодные годы карп в прудах совсем не размножается, и подготовленная к откладке икра перерождается и рассасывается. Такое же явление известно в отношении сазана в естественных водоемах, несомненно, то же можно наблюдать по отношению ко многим другим рыбам.
Определение температурного порога нереста
Одним из главнейших факторов внешней среды, сильно воздействующим на икрометание рыб, следует признать температуру воды. Нересту каждой рыбы свойственна определенная температура и минимальная температура колеблется мало. Для начала нереста карася нужна температура 17-18 °С, для уклей-15-16 °С, для леща-12-13 °С, для густеры-15- 16 °С (по наблюдениям П. А. Дрягина над рыбами озера Ильмень). Но это справедливо не для всяких водоемов.
В разгар нереста нужно провести круглосуточные (через каждый час) гидрометеорологические наблюдения. О важности микроклиматических данных для ихтиологических работ можно судить, например, по той сводке о нересте некоторых рыб, которая приведена А. В. Лукиным (1947). Лещ в средней Волге нерестует при температуре воды 10,0-13,5 °С, в дельте Волги-при 17-20, в низовьях Дона-при 23-24 °С. К сожалению, нет данных о методе определения начала нереста. Полагаю, что приведенные здесь данные не имеют нужной точности. Очень важно определять температурный порог начала нереста, для чего необходимо вести наблюдения за температурой воды и воздуха непосредственно на нерестилищах.
П. А. Дрягин (1952) пишет, что при определении начала нереста рыб очень важно учесть температурный порог, ниже которого у того или иного вида нерест не происходит. Его следует определять непосредственно на местах нереста в те часы, когда совершается нерест. Надо иметь в виду, что колебания температуры весной (иногда и осенью) в течение суток очень значительны и потому осредненные данные или случайные наблюдения могут дать совершенно ложные материалы о связи нереста с температурой воды.
В разных частях одного водоема или в разных водоемах даты начала нереста одного и того же вида рыб могут быть различными.
Понятно, что установление средних сроков и продолжительности нереста может быть получено только при многих и долголетних наблюдениях в различных водоемах. При наблюдениях за нерестом рыб нужно одновременно вести наблюдения и за соотношением полов. У рыб с порционным икрометанием нужно строго следить за началом и концом выметывания каждой порции икры.
Точно также необходимо было бы получить сведения о пороговых температурах, при которых в естественных условиях идет развитие икры. Можно рекомендовать постановку экспериментальных работ по выяснению этого существенного вопроса. И. Г. Юданов (1939) сообщил, что жизнеспособность икры сибирской ряпушки не исчезает даже в промерзших местах Обской губы.
Процессы откладывания и оплодотворения икры
Рыбы откладывают икру в разное время суток – утром, днем, ночью. Нужно отмечать такие часы для каждой наблюдаемой рыбы. М. И. Тихий (1939) наблюдал, что лещ (на озере Череменецком) нерестовую игру производит рано утром:«В ночь на 15 июня отметили легкие всплески, но в пятом часу утра было установлено бурление воды вне зоны тростника, т. е. на значительном расстоянии от берега». На этом участке мережи были полны текучими экземплярами леща. Приведенные строки показывают, что научное биологическое изучение над точным временем нереста леща было объединено с ловом. Такое сочетание дает более надежные данные, чем сообщения рыбаков.
Икрометание семги, по-видимому, происходит ночью. На беломорских реках ночью можно при помощи сильного света фонаря или пламени горящих смолевых щепок, пней наблюдать брачную игру рыб, откладывание и оплодотворение икры.У одних видов рыб икра рассеивается в толще воды или по дну, у других она прикрепляется к подводным предметам. Есть рыбы, которые устраивают гнезда, куда кладут икру. Гнезда для икры встречаются у лососей и у колюшки. Есть живородящие рыбы (в наших водах – бельдюга и морской окунь). С. Г. Соин (1947), изучив нерест европейского и амурского сомов, отмечает большое различие между ними: европейский сом откладывает икру в виде скоплений, и самец охраняет ее, амурский же сом икру разбрасывает на поверхности заросшего травой дна, и самец не охраняет ее. Эти виды сомов имеют и морфологические различия.
Характер нерестилищ
Насколько велико научно-хозяйственное значение исследований нереста и нерестилищ рыб можно судить хотя бы по факту длительной дискуссии, возникшей в отношении исследования нереста мурманской сельди. История этой дискуссии весьма поучительна. Написано много полемических статей, предложено несколько различных гипотез. Вопрос этот кратко и ясно изложен в статье Б. П. Мантейфеля и Ю. Ю. Марти – Исследование нереста мурманской сельди (1939).
Некоторые исследователи (А. И. Рабинерсон и др.) считали, что мурманская сельдь нерестится у берегов Норвегии, и молодь ее оттуда сплывает в Баренцево море. Другие (Е. К. Суворов, И. И. Месяцев и др.) полагали, что основная масса мурманской сельди нерестует в самом Баренцевом море. Эти диаметрально противоположные взгляды совершенно по-различному ориентировали сельдяное хозяйство Мурмана.
В 1938 г. Полярным институтом рыбного хозяйства (ПИНРО) были проведены на исследовательском судне «Персей» три экспедиции, в задачи которых входило отыскание и исследование нерестилищ и личинок сельди, добываемой в Баренцевом море. Исследовательские работы были поставлены очень широко и образцово. Участвовало 12 научных сотрудников и 19 лаборантов, которыми исполнен комплекс океанологических исследований, т. е. по гидрохимии, гидрологии, планктону, ихтиопланктону, бентосу, составу грунта и по ихтиологии. Методика исследований была разработана прекрасно. Рыба добывалась сетями и тралами. Планктон и ихтиопланктон собирали икорными сетками, рингтралом, мальковыми пелагическими тралами Петерсена и планктонными сетями Нансена. На каждой станции производилось обычно два лова: вертикальный, от дна до поверхности, и горизонтальный (на малом ходу судна) в слое воды от поверхности до 5 м. Рингтралом пользовались при косом лове, от дна до поверхности, с последующим поверхностным горизонтальным десятиминутным ловом. При увеличении размеров личинок сельди велись не только косые ловы, но также и серийные горизонтальные ловы несколькими рингтралами н икорными сетками, прикрепленными “к одному тросу на различных глубинах.
Микропланктон собирали батометром Нансена, а с поверхности моря зачерпывали воду ведром. Бентос и пробы грунта собирали дночерпателем Петерсена (площадь 0,25 я2), ножевой драгой, тралом Сигсби, сельдяным оттертралом и мальковым придонным бимтралом Расса.
Мы подробно остановились на методике исследования нереста и нерестилищ мурманской сельди, имея в виду, что этот инструктивный материал может быть использован при организации и других подобных исследований.
Экспедиции Полярного института установили, что основная масса сельди Баренцева моря нерестится на лофотенских нерестилищах, причем авторы замечают, что нельзя отрицать возможности нереста некоторого количества сельди и в Баренцевом море (в фиордах Мурманского побережья). «Несомненно», говорят авторы, «что с лофотенских нерестилищ молодь сельди ежегодно вносится в Баренцево море, что и обеспечивает устойчивость запасов сельди в этом море». Последующие работы ПИНРО осветили с достаточной полнотой биологию атлантическо-скандинавских сельдей (Марти, 1956).
Исследование нерестилищ должно быть поставлено в программу работ ихтиологов на всех исследуемых водоемах. Нужно знать места нереста рыб и изучить их с гидрологической и биологической стороны. Необходимо выяснить заполнение нерестилищ производителями, степень засоренности нерестилищ, установить возможность расчистки и вообще возможность улучшения нерестилищ.
Под нерестилищами понимаются точные (в определенных топографических границах) площади дна или поверхности водоема, где происходит икрометание того или иного вида рыб. Такие места обозначают на карте водоема и записывают длину и ширину (м), площадь (м2 или км2). Надо дать батиметрическую карту нерестилищ, основные сведения по газовому и солевому режиму, планктону и бентосу, грунтам, водной растительности, по составу рыбного населения, обитающего или встречающегося на нерестилище.
У морских рыб с демерсальной, опускающейся на дно, икрой нерестилища обычно велики, велики и нерестилища морских рыб с пелагической икрой. В том и другом случае, особенно во втором, чрезвычайно трудно очертить точные места нереста. А при пелагической икре, когда выметанная икра разносится петровым и постоянными течениями моря, нередко и невозможно точно назвать эти места. Тут приходится пользоваться косвенными показателями: нахождением скоплений плавающей икры и этапом ее развития. Такие опыты поисков рыбы при помощи скоплений плавающей икры нередко производятся. По плавающей икре проводились в 1947 г. довольно удачно поиски минтая (Theragra chalcogramma) в Корейском заливе Японского моря (Веденский,1949).
У рыб, откладывающих икру в реках и озерах, нерестилища обычно невелики и занимают они довольно определенные площади, размеры которых легко установить.
Определять площади нерестилищ необходимо для того, чтобы знать, насколько обеспечивают существующие нерестилища нормальное размножение рыб. У проходных рыб, например у дальневосточных лососей, наблюдается иногда такое переполнение нерестилищ производителями, что позднее пришедшим производителям нет свободного места для откладывания икры и они вынуждены раскапывать (разорять) гнезда с икрой, положенной ранее нерестившимися рыбами. Выброшенная из разоренных гнезд икра попадает в быстрый речной поток и погибает или уничтожается другими рыбами. Переполнение нерестилищ производителями отрицательно воздействует и на динамику размножения. Поэтому при исключительно большом подходе на нерестилища производителей численность молоди может быть меньше, чем при меньшем заполнении нерестилищ. Зная площадь нерестилищ, можно подсчитать, какое количество производителей наиболее целесообразно пропускать на нерестилища. На речных водоемах, например на дальневосточных реках, можно регулировать пропуск производителей.
В 1938 г. в Тихоокеанском научно-исследовательском институте рыбного хозяйства и океанографии проходила конференция по вопросам исследования дальневосточных лососей. На этой конференции были составлены (И. И. Кузнецовым, Ф. В. Крогиус, А. Я. Таранцом, А. Г. Смирновым и мною) программы и инструкции по изучению нерестилищ лососей. Наиболее существенные материалы мы приводим для использования при наблюдениях не только над дальневосточными лососями, но и над другими рыбами и в других водоемах. Программы включают и вопросы нереста рыб.
Паспортизация нерестилищ лососей
1. Сводка всех ранее полученных материалов по нерестилищам.
2. Сбор дополнительных (предварительных) данных о нерестовых водоемах (реках и ручьях), и о их расположении и размерах, о расположении и размерах нерестилищ, о количествах заходящей на эти нерестилища рыбы и сроках нереста.
3. Объезд нерестовых водоемов в момент нереста рыбы; буссольная или глазомерная съемка нерестилищ и описание их (грунт, глубина воды в момент посещения, геоморфологическое описание положения нерестилищ). По возможности надо определить, существует ли наличие или выход грунтовых вод. Определение типа нерестилищ (речной, ключевой, озерный). Определение количества рыб зашедших к моменту обследования на нерестилища, путем просчета на определенной нерестовой площади (на 20-25 м2) и их размещения на нерестилищах. Определение значения реки для того или другого вида рыб.
4. В случае посещения нерестового водоема во время нереста – определение тех нерестилищ, о которых получены сведения опросного порядка с учетом действительных возможностей нереста на данном участке, с подробным описанием нерестилищ. Учет гнезд с отложенной икрой.
5. Выявление возможностей увеличения нерестовой площади, путем простейших мелиоративных мероприятий. Определение условий захода рыбы на нерест, учет естественных препятствий для захода (пороги, водопады, мели), возможные колебания уровня.
6. При более детальных работах нужны гидрохимические анализы воды на нерестилищах для выяснения наличия грунтовых вод. Определение температуры и скорости течения.
7. Определение состояния нерестилищ (их засоренность, хищнический промысел).
8. Ориентировочное определение значения хищников на нерестилищах.
9. В случае повторного посещения нерестилищ проводить повторные контрольные работы.
Программа круглогодичных работ по исследованию нерестилищ дальневосточных лососей
Такая программа проводилась на р. Иски (в бассейне Амура) над дальневосточными лососями.
1. Метеорологические наблюдения.
2. Гидробиологическая характеристика бассейна реки. Фауна и флора в береговой и стрежневой частях реки. Бентос. Обрастание. Планктон.
3. Полуинструментальная съемка мест нереста лососевых в бассейне реки с целью учета площади и расположения нерестовых угодий.
4. Изучение биологии лососей (кеты и горбуши), идущих к местам нереста:
а) возрастной состав и вес;
б) половой состав;
в) степень зрелости и плодовитость;
г) изучение экологических условий подхода рыбы к нерестилищам для выявления влияния их на распределение лососей по различным нерестилищам.
5. Наблюдение над нерестом и нерестилищами во время нереста:
а) наблюдение над процессом нереста;
б) изучение сезонного распределения рыб по нерестилищам путем периодических обходов с нанесением нерестилищ на карту;
в) периодический, поштучный просчет нерестующей рыбы на выбранных типичных участках нерестилищ;
г) изучение погибшей рыбы: интенсивность гибели по времени, учет рыбы, погибшей до нереста;
д) раскопка бугров с заложенной икрой с целью установления характера закладки икры при различных условиях и количества икры в буграх;
е) изучение гидрологического и гидрохимического режима реки и нерестилищ во время нереста (определение t°, pH, C02, CaO, Mg, Si, N2Оз, Fe2O3, H2S).
6. Изучение развития икры и молоди для выявления причин колебания урожая молоди на нерестилищах в разные годы:
а) вскрытие бугров с целью установления интенсивности и причин гибели икры в течение сезона и в различных участках нерестилищ;
б) роль хищных рыб на нерестилищах лососей;
в) изучение гидрологических и гидрохимических условий развития икры и первых стадий развития молоди.
7. Изучение ската молоди:
а) мечение скатывающейся молоди вблизи нерестилищ;
б) контрольный лов молоди в устье реки с целью изучения интенсивности и характера ската, а также определения количества скатившейся молоди;
в) изучение гидрологических условий ската молоди (от нерестилищ до выхода в море).
Приведенные программы могут быть сокращены или расширены в зависимости от поставленных задач, но должны содержать следующие основные пункты:
1. Физико-географическое описание нерестилищ.
2. Гидрологическую и гидрохимическую характеристики.
3. Характеристику биоценозов (флора и фауна беспозвоночных и рыб) нерестилищ.
4. Наблюдения над нерестом (заполнение нерестилищ производителями), сроки нереста.
5. Наблюдение над развитием икры.
6. Наблюдения над выклюнувшейся молодью.
7. Сведения о лове рыбы на нерестилищах (техника лова н улов).
8. Сбор материалов по биологии производителей и молоди.
9. Состояние нерестилищ и мероприятия по улучшению нерестилищ.
Плотность кладок икры у лососей
Сведения по вскрытию гнезд лососей (за исключением тихоокеанских) немногочисленны. В отношении атлантических лососей, в частности северного лосося – семги, данные есть в работах М. П. Виролайнена (1946), П. И. Новикова (1947, 1948, 1949), В. С. Михина (1950), В. С. Михина и А. О. Шпайхера (1957), М. И. Владимирской (1957), Н. Д. Никифорова (1959). Бугры лососей можно вскрывать в два периода: вскоре после нереста и зимой, или ранней весной до схода льда. Вскрытия бугров позволяют выяснить: число гнезд в бугре, количество икры в гнезде и во всем бугре, число погибших икринок, глубину закладки икры, кучность икры в различных участках гнезда, расстояние отдельных гнезд друг от друга (в бугре).
Работа по вскрытию гнезд может дать более ценные результаты, если ее организовать на реке или речке, где представляются возможности вести наблюдения над семужьей молодью, над ее «урожайностью» в различные годы.
Если предполагается вскрывать гнезда зимой или ранней весной, следует осенью в период нереста отметить точно места, где происходила кладка икры. Для этого над гнездом вбивают колья в виде треножника и сверху прикрепляют груз – большие камни. Надо метить те гнезда, которые заложены ближе к берегу и ко времени наступления малого уровня воды в реке могут оказаться на более мелких местах. Наиболее опытные рыбаки хорошо знают такие места.
Зимой или ранней весной над помеченными осенью гнездами делают проруби. Если глубина воды над гнездами превышает 50 см, извлекать икру при быстром течении воды очень Трудно (видимость икринок уменьшается, икринки, попавшие в струю, быстро относятся течением). Чтобы уменьшить течение s- перед прорубью (вверх по течению), в 1,0-1,5 м от нее прорубают вторую удлиненной формы узкую прорубь, куда вставляют ветки и доски, которые образуют щит, ослабляющий течение до такой степени, что икринки, извлекаемые из гнезда, не уносятся течением.
Грунт с гнезда снимают обыкновенной железной лопатой с длинной ручкой. Икринки собирают маленькими сачками с мешочком удлиненной формы, сделанным из марли. Для фиксации икры можно пользоваться двухпроцентным раствором формалина.
При вскрытии гнезд записывают следующие данные: название нерестилища, где метились гнезда, расстояние от устья реки, толща льда, глубина воды над гнездом, скорость течения у гнезда и у поверхности льда. Из гидрохимических данных нужно определить: рН, кислород, окисляемость, железо.
Из гнезда берут пробу грунта весом не менее 1,5-2 кг для механического анализа. Наиболее удобно принять следующие размеры фракции для механического анализа грунта (Кленова, 1931).
|
Фракция |
Размер частиц, мм |
|
Сильт (пыль, ил) |
0,01-0,1 |
|
Песок |
0,1-1,0 |
|
крупный ……… средний ………… мелкий ……….… |
1,0-0,5 0,5-0,25 0,25-0,1 |
|
Гравий |
1,0-10,0 |
|
крупный ……… средний ……….. мелкий ………… |
5-10 2-5 1-2 |
|
Галька |
10,0-100,0 |
|
крупная ……….. средняя ………… мелкая ……….… |
50-100,0 20-50,0 10,0-20,0 |
|
Валуны Обломки более 10 см |
|
Одинаковая методика механического анализа грунта позволит иметь сравнительный материал.
Если при наступлении малого уровня воды в реке глубина бывает больше 50 см, то чтобы извлечь икру из гнезда, нужно применить водолазный костюм простейшего типа с аппаратом для дыхания атмосферным воздухом (без подкачки воздуха). Применение такого костюма дало хорошие результаты при гидробиологических исследованиях литоральной зоны озер (Моки-евский,1950).
Быстро развивающаяся техника подводных наблюдений при помощи акваланга открывает широкие перспективы и для исследования нерестилищ и размножения рыб.
Исследования озерных нерестилищ
П. А. Дрягин в своих работах приводит ценные методические советы по изучению озерных нерестилищ (1938). Полезные указания можно почерпнуть и из его работы «Половые циклы и нерест рыб (1949)». П. А. Дрягин делит нерестилища на 3 типа. Главнейшие сведения, которые нужно (по П. А. Дрягину) приводить при описании этих типов нерестилищ, следующие:
1. Мелкие нерестилища глубиной от 0 до 50 см, обычно 20-40 см. Подобные нерестилища свойственны плотве, язю, синцу, окуню, густере и некоторым другим рыбам. Они удобны для непосредственного осмотра и благодаря этому могут быть наиболее подробно описаны. Пробы грунта и растительности здесь можно брать руками. Для учета количества зарослей следует; закладывать пробные площадки. Стратификация, как правило, отсутствует, поэтому пробы для определения содержания газов берут в одном горизонте. Основные орудия, которыми приходится пользоваться при обследовании подобного типа нерестилищ, – скребок, железные грабли, сачок из частой сетки (лучше всего мельничный газ) и сетка Кори.
2. Нерестилища поверхностного слоя воды или «условно Мелкие» нерестилища. В таких нерестилищах плотва и лещ откладывают икру на субстрат, расположенный в поверхностных слоях воды при глубине места до 2 м и более. Так же, как и Предыдущий тип, эти нерестилища поддаются непосредственному обозрению, но учет растительности, обследование грунта и гидрохимические работы на них усложняются. Пробы грунта приходится брать или при помощи лота Воронкова, или дночерпателем. Определение кислорода, углекислоты и других гидрохимических элементов надо брать в нескольких горизонтах. Основные орудия для обследования – сетка Кори и сачок, последний лучше делать четырехугольным, определенной площади, тогда он будет пригоден и для количественных сборов.
3. Приглубые нерестилища (глубина свыше 70 см). На таких нерестилищах мечут икру снеток, ерш и другие рыбы с демерсальной икрой. В отличие от двух первых типов эти нерестилища не поддаются непосредственному наблюдению. Обнаружить их можно тралящим орудием лова подобно драге или маленькому бимтралу. В Карельском отделении ГосНИОРХ собирали икру на глубоководных с мягким грунтом нерестилищах ряпушки малым гидробиологическим тралом типа Сигсби. Трал Протягивали на определенном расстоянии (25-50 м), что позволило вести и количественный учет. Для учета смытых икринок. И плавающих личинок необходимо пользоваться сеткой Кори и различными планктонными сетками. Этот тип нерестилищ наиболее трудно обнаружить и изучить. Особенно трудно здесь проводить количественный учет. В некоторых случаях, когда нерестилище уже обнаружено и примерно определена его площадь, возможен количественный учет при помощи дночерпателей. Дночерпателем можно пользоваться только на нерестилищах с мягким грунтом и с не очень густой растительностью.
Важно учитывать требования рыб в отношении субстрата для кладки икры. С. Г. Крыжановский (1948) выделяет следующие экологические группы (по отношению к субстрату): 1) литофильная – рыбы, откладывающие икру на каменистом грунте; 2) фитофильная – размножающиеся на подводных растениях; 3) псаммофильная – размножающиеся на песчаном дне; 4) пресноводная пелагофильная – мечущие икру, развивающуюся в плавучем состоянии; 5) остракофильная – рыбы, откладывающие икру в жаберную полость двустворчатых моллюсков; 6) индифферентная – рыбы, откладывающие икру на различных субстратах; 7) гнездующая; 8) вынашивающая; 9) живородящая.
П. А. Дрягин (1952) советует различать такие основные биоэкологические группы рыб: 1) порционно нерестующие рыбы (яйцекладущие, живородящие и остракофильные): а) рыбы с круглогодичным нерестом; б) с сезонным порционным нерестом; нерестовый порог у пресноводных рыб этой группы обычно не ниже 16-20 °С; 2) единовременно нерестующие рыбы (яйцекладущие моноцикличные и полицикличные), нерестовый порог обычно ниже 10-12 °С.
Нерестилища снетка обычно приурочены к береговой линии, к поднятым частям берегового вала, с обилием корней осоки, выносами отмерших стеблей осоки и мха (Дрягин, 1949). Следует учитывать, что нерестилища подвержены всяким изменениям, они зарастают травой, заболачиваются, промерзают и т. д.
При акклиматизации рыб одним из главных условий, обеспечивающих успех акклиматизации, должны быть подходящие для размножения вводимой рыбы нерестилища.
Исследование всех условий размножения рыб, их нереста и нерестилищ должно занимать в ихтиологических работах первенствующее место, так как без этого невозможно правильно и полно осветить вопрос о состоянии запасов рыб. Если нерест промысловых рыб не обеспечен хорошими нерестилищами, то рыбные запасы не останутся на высоте. Обмеление водоема (естественное или искусственное) ухудшает или даже уничтожает нерестилища. Известны случаи, когда при таких условиях рыба вынуждена класть икру в такие места, где она не может нормально развиваться, или рыба вовсе не выметывает половых продуктов. Такое явление наблюдалось многими ихтиологами (Никольский и Морозова, 1946). Следовательно, ихтиолог, занимающийся изучением нереста рыб, должен помнить, что нерестом определяется состояние рыбных запасов и не ограничиваться только научной, биологической стороной этого явления. Практическая сторона исследований нереста и нерестилищ (найти меры, которые обеспечат производителям свободный проход на нерестилища, чтобы самые нерестилища соответствовали требованиям нерестующих рыб) нередко бывает более нужна, чем подробное (тоже необходимое) изучение всех процессов размножения.
Как пример, а таких примеров в русской литературе много, детального изучения нереста и нерестилищ рыб упомянем работу Г. Я. Дорошина и С. К. Троицкого (1949). В статье приводится характеристика условий размножения кубанской севрюги: сроки и места нереста, температура и высота уровня воды, скорость течения, мутность, хищные рыбы, поедающие икру, В распределение производителей на местах икрометания, количественный и качественный состав производителей (половой состав, размеры и вес производителей, возраст), эффективность запрета на лов севрюги, скат молоди как материал для суждения об эффективности икрометания севрюги, размеры (линейные и весовые) молоди. Такой способ исследования нереста и нереста лиц может служить программой для подобных исследовании и других рыб.
Орудия для сбора икры
Для сбора в море придонной и пелагической икры применяются специальные орудия. Мы описываем сеть для лова пелагической икры (по Рассу, 1939), которая используется и при работах на озерах. Сеть состоит из обруча, сетного мешка и стакана. Площадь зева равна 0,5 м2, общая длина 4 м.
![]() В собранном виде сеть представляет собой конус из шелкового газа, висящий вершиной вниз на обруче и поддерживаемый оттяжками(рис. 24). Вершиной конуса является стакан, основанием – входное отверстие сети. Обруч изготовляется из круглого железа диаметром 8-10 мм (или из железной или латунной трубки диаметром 12-15 мм). Диаметр обруча 80 см. Оттяжки (четыре) делаются и 6-миллиметрового фалшнура или лаглиня: верхние концы их привязываются к латунному кольцу (толщиной 8 мм, диаметром 4,5 см), которое служит для подвешивания сети к тросу. От верхнего кольца оттяжки прикрепляются к обручу сети, к колечкам пояска сети и к колечкам стакана. Нижние концы оттяжек (на 10 см ниже дна стакана) закрепляются на латунном кольце диаметром 4,5 см, Служащем для подвешивания груза при вертикальных ловах.
В собранном виде сеть представляет собой конус из шелкового газа, висящий вершиной вниз на обруче и поддерживаемый оттяжками(рис. 24). Вершиной конуса является стакан, основанием – входное отверстие сети. Обруч изготовляется из круглого железа диаметром 8-10 мм (или из железной или латунной трубки диаметром 12-15 мм). Диаметр обруча 80 см. Оттяжки (четыре) делаются и 6-миллиметрового фалшнура или лаглиня: верхние концы их привязываются к латунному кольцу (толщиной 8 мм, диаметром 4,5 см), которое служит для подвешивания сети к тросу. От верхнего кольца оттяжки прикрепляются к обручу сети, к колечкам пояска сети и к колечкам стакана. Нижние концы оттяжек (на 10 см ниже дна стакана) закрепляются на латунном кольце диаметром 4,5 см, Служащем для подвешивания груза при вертикальных ловах.
Рис. 24 Сеть для лова пелагической икры
(по Рассу, 1939) 1 – оттяжка; 2 – сито;
3-средний пояс; 4 – стаканный поясок;
5- стакан.
Сетной мешок состоит из холщовых частей и из усеченного конуса из шелкового газа№ 15 (15 ячей на 1 см) длиной 30 см по боку, 252 см по окружности основания (входного конца) и 25 см по окружности конца у стакана (верхушки). Сетной конус кроится и шьется, как для планктонных сеток (рис. 25), из двух полотнищ, с тем лишь отличием, что к каждому полотнищу приходится прикраивать еще по треугольнику, так как размеры полотнища превышают ширину куска газа.
Стакан (рис. 26 ) состоит из верхнего кольца, к которому подвешивается стаканный поясок сети, корпуса стакана и обжимного кольца. Стакан в собранном виде изготовляется из латуни.
Сетью производят вертикальные и горизонтальные ловы икринок.
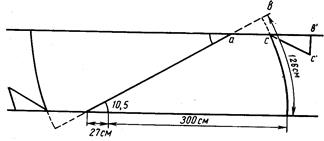
Рис. 25. Выкройка сетного конуса икорной сети (по Рассу, 1939).
Вертикальный лов делают во время стоянки судна на станции. Прицепленная к тросу чекелем или карабином сеть переносится за борт, и к нижнему кольцу ее прикрепляют груз. Груз осторожно (без рывка) переводят за борт, следя за тем, чтобы он все время висел на оттяжках. Сеть спускают до достижения обручем поверхности воды. Счетчик устанавливают на нуль и отпускают стопор на вьюшки, представляя сети идти вниз под влиянием собственной тяжести, изредка регулируя ее спуск стопором.
 Рис. 26. Стакан для икорной сети (по Рассу, 1939).
Рис. 26. Стакан для икорной сети (по Рассу, 1939).
Останавливают сеть, не доходя на 5 м до дна. ( В особенно мелководных районах кладут сетку на дно, опуская до посадки обруча.) Отмечают по угломеру отклонение троса от вертикали и выбирают сеть без остановки со скоростью 1 м/сек. Когда верхнее кольцо сети подойдет к счетчику, стопорят вьюшки и осторожно, поддерживая груз на оттяжках, переносят его и сеть на палубу, следя за тем, чтобы стакан сети все время сохранял вертикальное положение. Затем, расправив сеть, ополаскивают ее снаружи водой из ведра, отцепляют груз и снимают стакан. Низ стакана погружают до трети высоты (чтобы планктон всплыл) в ведро с водой, дно стакана прикрывают снизу ладонью, затем вынимают стакан и быстро выливают его содержимое (через воронку или непосредственно) в банку емкостью 300-500 см3.
Горизонтальный поверхностный лов производится на самом малом ходу судна. К обручу сети на случай ее обрыва прицепляют буйреп с буйком, причем или буйреп должен быть больше глубины или буек должен выдерживать сеть на плаву.
Прицепленную к тросу сеть переносят за борт и осторожно •спускают, пока стакан сети не коснется поверхности воды. Вследствие хода судна сеть оттягивается водой назад. Вытравив 30-35 м троса, стопорят вьюшку, производя лов в течение 10 мин от этого момента. Ход судна должен быть таким, чтобы верхний край обруча сети шел как раз под поверхностью воды. Чтобы не менять положения станции, судно должно описывать круг.
Через 10 мин после начала лова судно останавливают и немедленно выбирают сеть. Когда верхнее кольцо сети подойдет к счетчику, стопорят вьюшку и поднимают сеть на палубу, следя за тем, чтобы стакан сохранял вертикальное положение.
Затем сеть ополаскивают снаружи из ведра, снимают стакан и переносят сбор в банку, как описано выше для вертикального лова.
В озерах и реках сбор отложенной икры производится разнообразными способами и орудиями. Крупную икру, отложенную на мелких местах (икру семги), можно собрать в нерестовых буграх (гнездах), икру, рассеянную на песке (например, икру сигов) собирают дночерпателем и драгой, икру, высеянную на подводную растительность, собирают вместе с этой растительностью. Труднее собрать икру, прикрепленную к камням, – мелкие камни можно вытаскивать драгой, а с крупных камней можно счищать скребком. Для сборов икры, отложенной на грунт, пригоден, как уже упоминалось, гидробиологический трал Сигсби. Подробное описание такого трала есть в книге В. И. Жадина «методы гидробиологического исследования» (1960).
Способы учета отложенной и оплодотворенной икры
Методика изучения нерестилищ рыб требует дальнейшей разработки. Например, недостаточно установлены способы определения плотности засева нерестилищ икрой. Можно рекомендовать методы, которые применялись при работах Института биологии Карельского филиала Академии наук (Потапова, 1956). В небольшом озере (Миккельском) в продолжение трех лет учитывали нерестовые участки леща, участки, засеянные икрой. Оказалось, что в разные годы площади таких участков различны: в годы с высоким уровнем площади большие (до 450 м2), к маловодные годы-меньшие (до 150 м2), причем общая площадь, пригодная для засева лещовой икрой, определена в 20000 м2, т.е. икрой засевается менее 3% этой площади. Такой вывод, с одной стороны, указывает на недостаточное количество производителей леща, приходящих на нерестилища (из-за вылова их на путях хода на нерестилища); с другой – обосновывает мероприятия по пропуску производителей на нерестилища и даже по искусственному засеву лещовой икрой пустующих нерестилищ. Густоту засева нерестилищ икрой леща определяли на пробных площадках в 400 см2 (20Х20 см) на глубине 0,5 м; на нерестилищах с большей глубиной пробу брали граблями с площадки в 90 см2. Всю растительность с икрой клали в тазы с водой, и в береговой лаборатории просчитывали количество живой и мертвой икры, затем определяли среднюю густоту засева икрой на 1 см2 или на 1 м2 .
Интересная работа по учету кладок икры окуня в 5 окуневых и 5 окунево-плотвичных озерах Карельского перешейка выполнена в 1956-1961 гг. Л. А. Жаковым (Жаков, 1964).
Искусственные нерестилища
Искусственные нерестилища должны соответствовать условиям естественного размножения каждого вида рыб.
Устройство искусственных нерестилищ на озерах издавна применяется самими рыбаками с целью наиболее легкого способа вылова рыбы, скопившейся у таких нерестилищ – приманок. Описание устройств искусственных нерестилищ можно найти у многих авторов (Белый, 1951, 1952, 1961; Дрягин, 1938, 1939, 1940, 1952; Себенцов и Михеев, 1949; Вальмус и Правдин, 1923; Мороз, 1960 и др.).
П. В. Михеев (1953) описал плавучие нерестилища в дельте реки Дона для размножения судака и леща. К плавающим на поверхности воды деревянным рамам размером 6 м X 1 м через каждые 30-40 см прикрепляли поводки, длина которых равна глубине места установки нерестилищ. К каждому поводку через 30 см подвязывали венички, которые при помощи прикрепленного к концам поводков груза удерживались в воде в вертикальном положении. Рамы, лестницы плавучие, количество веничков и поводков могут быть различными, но рекомендуется верхние веники погружать на глубину не менее 25 см, на таком же расстоянии от дна должны отстоять нижние веники. Конструкция нерестилищ, материалы для веничков, места установок нерестилищ избираются в зависимости от исследуемых видов рыб. Эти требования нерестующих рыб ихтиолог должен выяснять, наблюдая за поведением пришедших на нерест рыб.
Во многих местах поставлен на очередь вопрос о снижении численности малоценных рыб (искусственные нерестилища могут быть одним из лучших способов создания больших скоплений таких рыб), которые легко обловить или уничтожить икру, отложенную на искусственных субстратах.
Великое преобразование природы, в том числе и водной природы нашей страны, поставило перед ихтиологами множество больших задач, от правильного разрешения которых зависит и повышение и устойчивость рыбных запасов рек и морей, где неизбежно нарушаются естественные нерестилища. Сооружение каналов, водохранилищ и гидроэлектростанций с грандиозными плотинами для одних рыб создают новые и лучшие нерестилища, для других ухудшают нерестилища. Поэтому изучение нереста и нерестилищ рыб в настоящее время приобретает совершенно исключительный интерес.
Глава VI
ИЗУЧЕНИЕ ЖИЗНЕННЫХ ЦИКЛОВ И МОЛОДИ РЫБ
Изучение жизненных циклов рыб
В изучении жизненных циклов рыб до сего времени не изжит целый ряд недостатков, которые в той или иной мере отрицательно влияют на разрешение этого важного вопроса. Из наиболее существенных недостатков можно указать:
а) разнобой в советской и зарубежной ихтиологической литературе в терминологии обозначений отдельных ступеней и моментов в онтогенезе рыб;
б) игнорирование изучения жизненного цикла рыб в целом;
в) недостаточный учет взаимозависимости между отдельными ступенями индивидуального развития и влияния их на жизненный цикл в целом.
Весьма полезные указания по упорядочению терминологии по изучению жизненных циклов рыб даны С. Г. Крыжановским (1956), который справедливо указывает на недопустимость применения терминов «предличинка» («прелярва»), «постлярва», «неоформившийся малек», «неоформившаяся личинка», «постэмбриональный период», а также термина «стадия» не только для обозначения каждого момента развития, но также для обозначения множества стадий развития.
В индивидуальном развитии рыб С. Г. Крыжановский предлагает различать эмбриональный, личиночный, мальковый и половозрелый периоды жизни с делением каждого периода на этапы, а этапы на стадии.
В соответствии с этой схемой подразделения жизненного цикла рыб С. Г. Крыжановский, Н. Н. Дислер, Е. Н. Смирнова (1953) устанавливают у судака 7 этапов в эмбриональном периоде развития, 5-в личиночном и 2 этапа-в мальковом периоде; Т. А. Детлаф и А. С. Гинзбург (1954) описывают в эмбриональном периоде развития осетровых рыб 36 стадий и 5 этапов; С. Г. Крыжановский (1956) устанавливает у сельдевых рыб 8 этапов в их эмбриональном периоде развития.
П. А. Дрягин (1961), принимая терминологию С. Г. Крыжановского за основу, рекомендует внести ряд уточнений в подразделениях жизненного цикла рыб и вместо употребляемого С. Г. Крыжановским термина «периоды» использовать для важнейших, наиболее ясно выраженных ступеней онтогенеза рыб термин «фазы» в том понимании их, как это было определено Фридрихом Энгельсом.
П. А. Дрягин в жизненном цикле рыб отмечает следующие фазы их индивидуального развития: эмбриональная-с момента оплодотворения яйца до вылупления личинки, личиночная-с момента вылупления личинки до начала образования непарных плавников, мальковая – с момента начала образования непарных плавников, до полного покрытия тела чешуей, неполовозрелости или юношеская – с полного покрытия тела чешуей до наступления III стадии зрелости, cозревания-от начала III до наступления V стадии зрелости у впервые нерестующих особей, половозрелости – он начала первого нереста и повторные половые циклы, старости-замедление, возможно и прекращение роста, ожирение, снижение плодовитости и половой активности и смерть.
В личиночной фазе особенное значение для выживаемости личинок имеют этапы перехода от эндогенного питания на смешанное и затем внешнее – экзогенное питание, в смысле обеспеченности соответствующими кормовыми организмами, а у некоторых видов карповых (например, у сазана) также наполнение плавательного пузыря атмосферным воздухом при соответствующих температурных условиях.
С. Г. Крыжановский личиночный период жизни определяет лишь с момента перехода личинок на смешанное питание, а жизнь личинок с эндогенным питанием относит к эмбриональному периоду, с чем едва ли можно согласиться.
В отношении изучения особенностей отдельных этапов развития получены многие ценные данные для эмбриональной, личиночной и мальковой фаз развития, но без достаточного учета отдаленных результатов воздействия разных факторов, вызывающих ускорение или замедление развития, а также влияния разнокачественности производителей и икры. Очень важно исследовать каждую фазу онтогенеза в морфологическом, экологическом, физиологическом и биохимическом отношении, а также поведение и распределение рыб после вылупления из икры.
Основная задача дальнейших исследований жизненных циклов рыб по П. А. Дрягину (1961) заключается, прежде всего, В поисках способов воздействия на разные и особенно на ранние фазы развития рыб, которые бы содействовали увеличению выхода ценных рыбных продуктов и ускорению оборота хозяйства. Иначе говоря, необходимо срочно заняться разработкой проблемы о направленном индивидуальном воздействии на развитие рыб в целях создания изобилия рыбных продуктов, помня при этом, что особенно чувствительны и отзывчивы рыбы на ранних стадиях развития и что реактивность организмов с возрастом падает. Так, в отношении растений И. В. Мичуриным (1948) установлено: «Всякое растение имеет способность изменяться в своем строении, приспособляясь к условиям новой среды лишь в молодом возрасте, и эта способность проявляется, начиная с первых дней после выхода из семени в большей мере, с течением времени постепенно слабеет и затем совершенно исчезает при полной возмужалости деревьев».
Исследований по рыбам в этом направлении пока ведется очень мало и возникает необходимость в проведении больших экспериментальных работ, а также в расширении и углублении исследований жизненных циклов разных видов рыб и их экологических форм, а также отдельных популяций и даже отдельных генераций, особенно у порционно нерестующих рыб при различных условиях обитания.
Для экспериментальных работ можно рекомендовать изучение следующих вопросов:
1. Влияние на потомство разнокачественности производителей (по возрасту, размерам, упитанности и другим показателям), а также разнокачественности икринок и спермы.
2. Эффективность оплодотворения в разных условиях обитания при естественном ходе созревания половых продуктов и при гипофизарной инъекции.
3. Влияние ускорения и замедления развития на разных эта пах и стадиях в эмбриональном периоде на выживаемость и жизнедеятельность эмбрионов и на последующие фазы онтогенеза.
4. Влияние ускорения и замедления развития и роста на разных этапах и стадиях личиночной, мальковой и юношеской фаз на жизнедеятельность, выживаемость, время наступления половозрелости, продолжительность жизни, количество повторных созреваний.
5. Ведущие факторы среды, воздействующие на развитие и рост рыб (температура, газовый режим, питание и т. д.) на разных этапах и фазах онтогенеза и их оптимальные показатели.
Для проведения исследований жизненных циклов рыб в природных условиях полезно наметить следующие вопросы:
1. Влияние плотности стад рыб на развитие, рост, сроки созревания, возрастной состав и продолжительность жизни, а так же на образование медленнорастущих и быстрорастущих популяций.
2. Факторы, определяющие биологическую неоднородность у разных видов рыб и их дифференциацию на экологические группы и сезонные расы.
3. Оптимальные условия для развития и роста основных видов промысловых рыб и их отдельных стад.
4. Биологические и рыбохозяйственные показатели лучших о промысловом отношении популяций отдельных видов рыб.
5. Сравнительные показатели по отдельным фазам жизненного цикла ценных видов рыб при культурном и стихийном (диком) ведении хозяйства.
Исследования по направленному преобразованию жизненных циклов рыб важны не только для практических целей, но и для вскрытия закономерностей взаимоотношения организм – среда М для познания причин и процессов формообразования и видообразования у рыб.
Изучение молоди рыб
Количество и качество молоди служат в большей степени, чем количество икры, индикатором состояния запасов всего ста да данного вида рыб. Обилие хорошо растущей молоди обеспечивает большее количество взрослых рыб и притом рыб с хорошим темпом роста, поскольку хороший рост молоди в первые годы благотворно влияет на рост рыбы в последующее время. Однако эта закономерность проявляется только при известных условиях. Рыбоводные опыты наглядно показывают, что даже при самых оптимальных условиях питания молоди количество ее в водоеме нередко бывает крайне ограничено. Какая же здесь причина?
При работах по выращиванию сазана в пойменных водоемах мы наблюдали случай, когда отсаженный в очень кормное, но не освобожденное от хищников озеро малек сазана (80 тыс. шт.) был очень быстро уничтожен молодыми судаками и щуками. При осеннем облове сазана почти не было. За этим опытом велись тщательные наблюдения. У вскрытых судаков и щук в желудках было много сазанчиков. Такое же явление, несомненно, бывает и во всех подобных водоемах. Степень выедания молоди хищными рыбами очевидна, но пока в рыбоводной литературе имеются лишь общие рассуждения по этому поводу, они не облечены в цифровую форму и поэтому мало убедительны. Тут необходим серьезный эксперимент. В приведенном выше опыте есть также крупнейший дефект: мы учли в точных цифрах жертву (мальков сазана), но не учли в цифрах истребителей (суда ков и щук). При правильном эксперименте необходимо учитывать и мальков и хищников, и только после этого можно вычислить коэффициент выживаемости молоди рыб.
При огромном выходе молоди может возникнуть недостаток в пище, способный привести эту молодь к полной дистрофии и даже к массовой гибели. Это также часто наблюдается в пойменных водоемах. Слабый рост молоди лососей в речной период жизни объясняют недостатком корма. Причины этого не ограничиваются вышеуказанными факторами (отсутствием или изобилием хищных рыб или пищи); хищники из мира беспозвоночных, паразиты, болезни – все это находится в теснейшей связи со стадом рыб и прежде всего со стадом молоди рыб, и все требует особо тщательных наблюдений и учета.
Изучение биологии молоди необходимо и для мероприятий по воспроизводству рыб. Непосредственных наблюдений над молодью рыб пока еще немного. О росте ее мы больше судим по структуре чешуи, чем по наблюдениям в природе. Следует ставить специальные работы (темы) по наблюдениям над жизнью молоди и сопровождать эти наблюдения работами экологического порядка (гидробиология и гидрология должны здесь занять главнейшие места). Современное рыбоводство требует выращивания молоди разводимых рыб (прежде чем выпускать их в промысловый водоем) и успех этого будет тем эффективнее, чем больше и полнее мы будем знать биологию молоди.
Большая проблема – изучение систематики молоди рыб. Нельзя изучать молодь лосося, не умея отличить ее от молоди форели.
Теперь, когда вопрос видообразования трактуется столь глубоко, что различие или сходство признаков живых организмов обнаруживается не только у взрослых и у молодых организмов, но даже во внутренней структуре их клеток и тканей, изучение молоди рыб нужно начинать с развития эмбриона, личинки, малька.
При изучении признаков эмбриона и личинок рыб необходим микроскоп.
Изучение молоди рыб давно привлекало внимание ихтиологов. Среди русских ихтиологов в 1904 г. была создана специальная комиссия в составе Н. М. Книповича, И. Н. Арнольда и Н. А. Бородина, которая разработала и предложила инструкцию по собиранию и изучению молоди рыб. Авторы инструкции рекомендовали вести аквариумные наблюдения над изменением «внешнего облика малька при его развитии» и указывали, как на наиболее верное средство видового распознавания мальков – применение искусственного оплодотворения и выведение молоди из икры.
Систематическая и серьезная работа по исследованию молоди рыб у нас начата в Астраханской ихтиологической лаборатории В. И. Казанским еще до революции. С 1915 г. В. И. Казанский опубликовал несколько работ по молоди рыб; особенно привлекают внимание его работы: «Материалы по развитию и систематике личинок карповых рыб» (1915) и «Этюды по морфологии и биологии личинок рыб нижней Волги» (1925).
А. М. Шуколюков (1932) привел морфологические и биологические данные по молоди рыб р. Урала.
Многое сделано по методике исследования молоди рыб Т. С. Рассом, которым составлены инструкции таких исследований (1935) и даны указания по орудиям и способам лова молоди.
Т. С. Расе различал следующие фазы развития рыб: а) личинки-«характеризуется наличием желточного мешка»; б) неоформившегося малька-«характеризуется отсутствием желточного мешка и отсутствием лучей в спинном и анальном плавниках», эта стадия длится от момента всасывания желточного мешка до появления лучей в спинном и анальном плавниках: в) оформившегося малька-характеризуется наличием лучей, до появления чешуи.
Рис. 27. Фазы развития трески (по Рассу, 1939):
И-икринка; Л-личинка; MI-малек неоформившийся, МII-малек оформившийся.
Описанные Т. С. Рассом фазы изображены на рис. 27. Позднее (в 1946 г.) Расе предложил другую номенклатуру индивидуального развития рыб. Он различал две категории ступеней развития: фазы и стадии. Четыре основные фазы: икринка (ovum), предличинка (praelarva) или личинка с желточным мешком (Правильнее фазу «предличинка» называть «личинка с эндогенным питанием».), личинка (larva) и малек (juvenis), обычно сеголеток. Эти четыре фазы -существенно отличаются между собой морфологически и экологически». «Фаза личинки длится от моментарезорбции желтка до окончания метаморфоза, обычно совпадающего с появлением чешуи на боках тела». Фаза малька или молоди начинается после окончания метаморфоза и длится до наступления половозрелости.
Фазы развития рыб Расе подразделяет на стадии.
В работе М. П. Рыженко (1939) имеется ряд полезных сведений по изучению личиночных и мальковых стадий мурманской сельди. Пелагических личинок сельди ловили икряными (диаметр зева 1 м) и планктонными (диаметр зева 0,5 м) сетями путем вертикальных (от дна до поверхности) и горизонтальных обловов. Икру и придонных личинок ловили мальковым тралом Расса (см. ниже). Применяли драги с газовой сеткой внутри и дночерпатели. В заливах мальков ловили тяглыми мальковыми неводами с ячеёй 4-5 мм и наживочными неводами с такими же размерами ячей в кутке. В открытых частях моря употребляли 6-футовый оттертрал с ячеёй 8-10 мм, пользовались также рингтралами с ячеёй 4-5 и 6-8 мм, с диаметром круга 1,5 м.
Н. В. Европейцева (1949а), занимаясь морфогенезом молоди трех форм ладожских сигов (волховского, свирского и лудоги), выращиваемых в прудах, различает следующие стадии развития (по морфологическим признакам) личинок сигов (с момента выклева до появления чешуи):
I. Стадия желточного мешка (рис. 28, а) – с момента выклевывания и до полной резорбции желточного мешка. Характеризуется наличием пассивного питания.
II. Стадия начала формирования хвостового плавника (рис. 28,6). Начинающий формироваться хвостовой плавник представлен скоплением мезенхимных элементов вокруг конца хорды или первыми гипуралиа и продолжается до начала загибания вверх уростиля.
III. Стадия образования симметричного хвостового плавника и окончательного формирования лучей во всех плавниках (рис. 28,в).
IV. Стадия предмальковая: после сформирования плавниковых лучей и до появления чешуи (рис. 28, г).
Номенклатура стадий личиночного периода, предлагаемая Н. В. Европейцевой, немного отличается от номенклатуры Т. С. Расса. Поэтому при подобных работах нужно ссылаться на автора, методика которого принимается. Европейцева подробно описывает каждую из четырех принимаемых ею стадий развития молоди сигов. Работа Н. В. Европейцевой снабжена хорошими рисунками личинок сигов. Этими рисунками можно пользоваться как определителем для распознавания нескольких форм ладожских сигов. У Н. В. Европейцевой есть работа и по личинкам окуневых (19496). Подобные исследования необходимо продолжить и по другим промысловым рыбам.
Об этапах развития молоди рыб можно найти материалы в работах В. В. Васнецова (1947) и других авторов. М. Ф. Вернидуб (1950) проследила темп роста личинок плотвы в зависимости от развития различных систем органов и пришла к выводу, что рост этих личинок проходит этапами. Автор различает 4 качественно различных этапа:

Рис. 28. Стадии развития личинок сига (по Европейцевой, 1949):
а-стадия желточного мешка; б-стадия начала формирования хвостового плавника; в – стадия формирования симметричного хвостового плавника; г – стадия предмальковая.
I-при длине личинки 8-11,5 мм, II-при длине 14,5- 16 мм. III-при длине 19-22 мм, IV-при длине 26-29 мм. Каждый этап автор характеризует и физиологически.
Много полезного по изучению морфологии молоди рыб содержится в работах Б. В. Веригина (1950), Я. И. Гинзбурга 1 (1950) и М. М. Мешкова (1951). Работы снабжены рисунками. Успех выращивания на рыбоводных заводах молоди ценных промысловых рыб всецело определяется степенью изучения биологии молоди этих рыб в естественных условиях. Знание биологии и экологии молоди рыб, начиная с эмбриона, абсолютно необходимо для освещения вопросов по динамике численности рыб. В первую очередь этими причинами следует объяснить небывалый рост литературы по молоди рыб. И в учебниках (например, у Б. И. Черфаса, 1956) и особенно в специальных статьях последнего времени можно найти необходимые сведения о молоди рыб и о методике исследования молоди. В этом отношении необходимым пособием, кроме названных выше, может служить 11-й том Трудов Амурской ихтиологической экспедиции 1945-1949 гг. Главнейшей статьей в этом томе служит большая статья С. Г. Крыжановского, А. И. Смирнова, С. Г. Соина-Материалы по развитию рыб р. Амура (1951). В статье содержатся многочисленные данные по морфологии, анатомии и биологии эмбрионов, личинок и мальков рыб, а также по икре и нересту многих рыб. Статья снабжена многочисленными рисунками, облегчающими видовое распознание личинок.
Методический интерес представляет статья Е. С. Кожиной (1956) по наблюдениям над ранними стадиями жизни леща. Автор наблюдал эмбриональное развитие леща в естественных и лабораторных условиях и проследил развитие личинок и мальков от трех нерестовых сезонов (1952, 1953, 1954 гг.), что обеспечило получение более надежных выводов. При понижении температуры воды ниже 11° лещ приостанавливал нерест и возобновлял его при 12 °С. Икра леща способна развиваться при значительном снижении содержания кислорода в воде (до 54% О2); диаметр икринки в среднем 1,99 мм. Подробно описано развитие оплодотворенной икры. Такие же подробные наблюдения у Е. С. Кожиной и над личинками и мальками леща. В книге приведены соответствующие рисунки.
Для изучения морфологических признаков часть мальков отбирается для подробных измерений. Рекомендуется взять 30 мальков в первый момент после всасывания желточного мешка, через 5 дней снова 30 рыбок и еще одну такую же пробу через 5 дней после второй, затем 3 пробы по 30 мальков через каждые 10 дней, потом 3 таких же пробы через каждые 15 дней и далее брать молодь через каждый месяц также по 30 штук. Здесь указано минимальное число рыб (30), которое можно будет обрабатывать обычными вариационно-статистическими методами.
Если исследователь сможет продолжить сбор месячных проб до той поры, когда рыба станет половозрелой (у многих рыб это наступает на 3-5-м годах жизни), то, несомненно, будет получен ценнейший научный материал, обработка которого углубит познания проблемы- морфологии вида.
Сбор материала по морфологическим признакам мальков следует сочетать с этапами развития, но для огромного большинства рыб такие этапы еще не окончательно установлены, а поэтому пока приходится группировать сборы мальков совершенно искусственно (через 5, 10, 15 и 30 дней после рассасывания желточного мешочка).
В. И. Казанский советует при сборе материала по молоди карповых рыб останавливаться нанести стадиях развития молоди-в стадии выклевывания, в стадии 3, 7, 14 и 21 дня и в стадии месячного возраста. Так как в эти моменты представлены последовательно все особенности строения личинок, то они могут служить для характеристики развития отдельных видов.
Для рыб других семейств, очевидно, могут быть другие сроки.
Следует признать, что изучение морфологических признаков молоди рыб представляет исключительные трудности, так как этим вопросом ихтиологи занимались очень мало, и до сих пор еще нет разработанных и общепринятых методов. Между тем, правильное распознавание молоди рыб, начиная с личиночной фазы, помимо систематической группировки молоди, может пролить свет на самые главные стороны биологии рыб: на установление времени и мест нереста, на миграции мальков и т. п. В этом отношении прав Т. С. Расе (1933), когда говорит: «Отыскание первых стадий имеет значение для выяснения условий нереста и определения нерестилищ, так как нахождение стадии неоплодотворенной икры (нулевой) и стадии дробления (первой) указывает на непосредственную близость мест нереста. Сбор (икры и личинок) заносится на карточку с обозначением гидрологических и метеорологических данных, что позволяет выяснить условия, сопутствовавшие сбору, и в случае преобладания в сборе первых стадий развития подметить условия нереста рыб».
И. И. Казанова и Н. А. Халдинова (1940), Т. А. Перцева (1940) на основании распределения икры и личинок каспийских сельдей установили места и условия нереста этих рыб в волжской дельте. В работе даются морфологические признаки икринок, таблица для определения икринок, указаны места распределения икринок, личинок, мальков сельдей.
Из морфологических признаков сеголетков нужно учитывать:число чешуи в боковой линии, или число поперечных рядов чешуи, число жаберных тычинок, ветвистых лучей в спинном и анальном плавниках, длину тела, диаметр глаза, длину головы, высоту тела, антедорсальное расстояние, длину хвостового стебля, длину и высоту спинного плавника, длину грудного плавника, верхней и нижней лопастей хвостового плавника. Названные признаки, по-видимому, имеют (одни большую, другие малую) зависимость от возраста рыбы, но этими признаками не покрываются все возрастные изменения морфологических признаков. Много полезного для изучения молоди рыб ихтиолог получит из книги Н. Л. Чугунова (1928). В. И. Казанский (1924) дал простую схему для определения мальков линя, карася, сазана, чехони, красноперки, уклей, густеры, белоглазки, леща, воблы, синца, рыбца, кутума, язя, подуста и жереха. Пользуясь схемой Казанского, можно определить названных рыб, начиная со стадии выхода из икры и до 10-дневного возраста. В основу схемы положены отличительные признаки пигментации.
При исследовании мальков рыб необходимо принимать во внимание рисунок (окраску) чешуйного покрова. Этот рисунок у молоди нередко не соответствует рисунку взрослых рыб. Например, молодь горбуши темных пятен не имеет, а у взрослой они есть; молодь нерки с пятнами, взрослая без пятен. В работах американских и русских авторов есть определительная таблица и рисунки молоди тихоокеанских лососей (Лагунов, 1939).
Существуют составленные ВНИРО таблицы для определения икры и личинок каспийских сельдей. А. И. Дехтярева (1940) дала подробные опознавательные признаки сеголетков каспийских сельдей. М. М. Мешковым (1951) дан определитель личинок карповых рыб.
Если мальки не могут быть определены, то производится только просчет их и запись в журнале, а затем их фиксируют слабым раствором формалина (1 : 19) в отдельной банке и снабжают этикеткой. Из формалина мальков следует потом переложить в спирт 60-70°.
Значение пигментации для распознавания сельди черноспинки особенно хорошо описано Б. И. Диксоном (1905). Прежде всего пигмент выражен в виде резкого пунктира из коротких, совершенно прямых черных штрихов, напоминающих знаки на телеграфной ленте. Затем он появляется приблизительно на месте спайки задних жаберных дужек на глоточном своде и идет в виде пунктира до внешнего края последней (задней) жаберной дуги; отсюда эта линия пигментных черточек раздваивается и идет вдоль всего тела по бокам брюшной части, по верхней границе просвечивающего кишечника и прерывается, не доходя до места образования брюшных плавников.
При изучении молоди неизбежно приходится проводить опыты по искусственному оплодотворению икры и по выращиванию личинок и мальков исследуемых рыб, потому что определение личинок без контрольных сравнительных коллекций личинок, отнесенных к точно установленным видам, представляет исключительные трудности. Искусственно выведенные личинки и могут служить такими контрольными коллекциями.
Искусственное оплодотворение икры и выведение личинок трудностей не представляет, и методика этой работы подробно описывается в многочисленных руководствах по рыбоводству. Однако более полезно для данного случая использовать указания Н. И. Николюкина, изложенные им в статье «К методике искусственного рыборазведения» (1935).
При искусственном выводе личинок рыб нужно иметь в виду, что оплодотворение икры бывает ударным в том случае, когда икра берется вполне зрелой и здоровой. А. И. Мищенко (1938), инкубируя икру камбалы, нашел, что лучшие результаты получаются, когда икру оплодотворяют немедленно после подъема рыбы (из трала), и когда рыба имеет текучие половые продукты. Оплодотворение велось в банках (1-3 л) с водой, куда осторожно прибавлялись молоки 1-2 самцов, затем тем же Способом, по стенке или по бумажке, добавляли зрелую икру и, наконец, опять молоки. После этого банку слегка покачивали для лучшего перемешивания икры со сперматозоидами и завязывали марлей. Однако развитие икры камбалы (икра камбалы пелагическая) при этом способе шло недостаточно хорошо. ‘: Выход личинок из икры начал наблюдаться на 4-й и особенно на 5-й день после оплодотворения. Смена воды в банках производилась простым переливанием через марлю или газ или резиновой трубкой, конец которой обвязывали этим же материалом.
В статье Мищенко приводятся рисунки двухдневных личинок (Limanda aspera и L. herzensteini). У личинок тело совершенно симметричное.
L. aspera:
Желточный мешочек удлиненный; отношение его высоты к длине равно 1:2.
Пигмент расположен на спинной части тела и голове в виде мелких разбросанных точек и т.д.
L. herzensteini:
Желточный мешочек овальный, отношение высоты к длине равно около 3 :4.
Пигмент расположен четырьмя скоплениями, из них три в постанальной части и одно в анальной и т. д.
Образцовое описание искусственного выведения личинок беломорской наваги дано Н. А. Халдиновой (1936). Опыты ставились в лабораторных условиях и в море. Инкубация икры лучше шла в глубокой фарфоровой тарелке и в часовых стеклах, где яйца не скатывались и лежали одним слоем. Заболевание сапролегнией предупреждалось 60-минутным выдерживанием икры в 2%-ном растворе NaCl в морской воде, как рекомендует Е. Г. Шерешевская (1932). Для нормального развития икринок наваги нужно проводить опыты при температуре воды от минус 1 до плюс 3°С.
Порог или рубеж развития, т.е. минимальная температура, ниже которой эмбрион развиваться не может, определяется формулой Рейбища (Reibisch, 1902) и для икринок наваги он равен минус 2,3 °С. Получив этот порог, Н. А. Халдинова вычислила гумму тепла, или число градусо-дней, потребных для развития икры, что равно 163,5-164,5 градусо-дней. Автору удалось проследить эмбриональное развитие и развитие личинок наваги, и -это дало возможность установить отличительные признаки личинок наваги от родственных ей личинок трески и сайки.
Формула Рейбиша, которой можно пользоваться при определении порога развития рыбы, имеет такой вид (по Черфасу, 1956):
![]()
где х – температурный порог;
t1 и t2– средние температуры в период развития;
п1 и п2,-длительность периода развития в днях или часах от момента оплодотворения икры до момента выклевывания личинок.
Сумму тепла, потребного для развития икры, или градусо-дни находят, умножая число дней, в продолжении которых шло развитие икры, на средние показатели температур, при которых шло развитие.
Подробные пояснения формулы Рейбиша и других вычислений, относящихся к методике изучения развития икры, в общедоступной форме изложены Б. И. Черфасом в его книге-Рыбоводство в естественных водоемах (1956). При экспериментальных работах по выведению молоди рыб ихтиолог должен ознакомиться с этой методикой.
Улов молоди тщательно разбирают и просчитывают по видам мальков. Если улов невелик (до 250-300 рыб), то весь улов промеряют с помощью штангенциркуля, обыкновенного школьного циркуля или (еще удобнее) специальной линейкой с точностью до 1 мм (определяют длины тела: аb, ас, ad и od). Взвешивают каждого малька.
Если улов велик, то из него предварительно отбирают всех редко встречающихся мальков, подлежащих измерению. Затем из основной массы улова отбирают пробу в количестве 180-200 мальков. Пробу берут объемным методом (половина, четверть и т.д. всего улова), сортируют по видам, измеряют и по группам (для каждого вида) взвешивают. При групповом взвешивании должен быть точный штучный просчет для последующего вычисления общего количества и среднего веса мальков.
Все эти данные записывают в журнал измерений мальков с указанием места и даты лова.
В дневник записывают общие наблюдения над ходом данных работ с краткой характеристикой распределения мальков, .различия между дневными и ночными уловами и т.д.
В журнале по изучению молоди рыб необходимо давать описание места, где и в какое время держатся мальки.
Измерение молоди рыб более удобно производить в лабораторной обстановке. Личинок и мальков на ранних стадиях измеряют с помощью окуляр-микрометра.
Места нахождения личинок и мальков отдельных видов рыб различны, и отсутствие мальков в исследовательских ловушках иногда не может служить доказательством отсутствия мальков в водоеме. Необходимо изучить поведение мальков в связи с условиями их обитания. Относительно мальков некоторых рыб известно, что они пасутся в самой береговой зоне, в тихой воде (карповые), других мальков можно найти только среди камней, в затемненных местах (лососи), у третьих мальки избегают темноты (плотва, лещ и др.).
Количество молоди рыб, а также и выживаемость отложенной в водоемы икры во многих случаях определяется степенью чистоты или загрязненности водоемов. Сточные воды целлюлозно-бумажных фабрик отравляют тот участок реки или озера. Куда они спускаются. Это необходимо учитывать при исследовательских работах и принимать соответствующие меры.
Орудия лова молоди рыб
Лов личинок требует специальных орудий.
Личинок, у которых желточный мешок уже всосался, а также мальков ловят сеткой Кори (рис. 29) из шелковой материи или канвы с ячеёй 2,5 мм, длиной-150 см, высотой 60 см.
![]() В конце сетки прикрепляется металлический стакан с шелковым газом. Кроме сетки Кори допускается применение мальковых кругов (рис. 30) диаметром не менее 1 м из указанного выше материала и салазочных Тралов (трал Сигсби).
В конце сетки прикрепляется металлический стакан с шелковым газом. Кроме сетки Кори допускается применение мальковых кругов (рис. 30) диаметром не менее 1 м из указанного выше материала и салазочных Тралов (трал Сигсби).
Сетку Кори выставляют на течение на определенное время: 5, 10 и 15 мин (в зависимости от обилия мальков), считая достаточным улов 150- 200 шт. Сетку спускают под поверхность воды так, чтобы верхняя подбора была на поверхности, причем нижнюю подбору следует пригрузить свинцом или ташами (каменными грузиками).
Употребляют и гельголандскую сеть с распорной доской и пелагический оттертрал. Описание гельголандской сетки имеется в книге Д. Джонетона-Условия жизни в море (перевод Н. М. Книповича, 1919).
Методика лова мальков рыб хорошо разработана и описана во многих статьях проф. Т. С. Рассом. В специальной инструкции Т. С. Расе (1939, 1958) дает подробные описания и рисунки малькового оттертрала, малькового бимтрала, рингтрала, малькового трала Петерсена и мальковой лампары. Подробности о названных мальковых ловушках (детали и способы изготовления) излагаются по инструкции Расса.

Рис. 31. Кройка пятиметрового малькового оттертрала (по Рассу, 1939):
а-сквер; б-нижние крылья; в-передняя часть матицы; г-задняя часть матицы;
д-мотня; е-клапан.
Кильковый (мальковый) оттертрал-основное орудие лова придонных сеголетков рыб. Сквер изготовляется из хлопчатобумажной дели № 20/9, крылья и передняя часть матицы -из дели с ячеёй в 12 мм от узла до узла, задняя часть матицы – 8 мм и мотня – 5 мм. Длина верхней подборы 5 м, нижней – 8 м, грунтропа – 6,9 м. Длина трала от середины верхней подборы до начала мотни – 6,5 м. На грунтроп прицепляют 4-6 кг железной цепи. Кройка трала показана на рис. 31. Размеры досок для трала: длина 70 см, высота 40 см. Устройство досок показано на рис. 32.
Перед пуском трала завязывают мотню и к ней подвязывают буй с буйрепом в 1,5 раза больше глубины (в особо мелководных водоемах-в Северном Каспии, Азовском море).
Уздечку трала прицепляют через вертлуг и чекель к концу ваера.
Пуск трала производится на самом малом ходу. Выбрасывают буй, мотню и сеть, затем спускают обе доски. Слегка задерживают ваера, давая разойтись доскам. Когда доски разошлись, травят ваер, пока трал не сядет на дно; вытравляют ваера на длину, от 1,5 до 5 глубин (в мелких районах). Время лова считается от момента посадки трала на дно. Трал тянут на самом малом или малом ходу (1-2 мили/ч} и все время следят по подергиванию ваера за тем, чтобы трал шел по дну. По истечении времени (тянут 15 мин) стопорят машину и выбирают трал лебедкой или руками.
Рис. 32. Устройство досок малькового оттертрала (по Рассу, 1939).
Рис. 33. Мальковый бимтрал Расса (по Рассу, 1939).
Мальковый бимтрал (конструкция Расса) является основным орудием для лова донно-пелагических икринок и личинок рыб. Состоит из следующих частей (рис. 33): основной рамы, башмаков, бима и сети трала. Основная рама сделана из полосового железа 30Х4 мм и 3,5-дюймовой газовой трубы (наружный диаметр 88 мм, внутренний-80 мм). Внутренние размеры рамы: длина 95 см, ширина 32 см; длина ножек (через которые проходит ось трубы) 87 мм. Длина трубы 925 мм.
Башмаки трала представляют собою полозья из дугообразно изогнутого полосового железа 80Х3 мм, стянутые по заднему краю перекладиной из полосового железа 30Х4 мм. Бим-круглое бревно с четырехгранными концами. Длина его 112 см, диаметр 12 см.
Рис. 34. Рама трала Расса (по Рассу, 1&39):
1-башмак; 2-верх рамы; 3-низ рамы; 4- бока рамы; 5-скоба; 6-втулка для оси трубы;
7-труба; 8-ось трубы; 9-петля для уздечки; 10-рама для сети.
Сеть трала надета на четырехугольную железную раму. Сверху и снизу к сети крепятся полотнища фартуков из холста или из грубой дели.
При сборке трала пространство между бимом, верхними краями башмаков и основной рамы, а также просвет башмаков обтягивают 10-15-миллиметровой делью, образующей сквер и крылья трала.
Для работы с малых судов и баркасов удобнее применять. малую модель трала Расса, отличающуюся мелкими размерами; деталей и упрощением конструкции основной рамы. Размеры и устройство деталей показано на рисунках 34 и 35.
Два башмака или полоза изготовляются из полосового железа 60х3 мм, длиной 117 см. Вдоль внутреннего края идут отверстия диаметром 88 мм, отстоящие (центры отверстий) от края на 10 мм и друг от друга на 50 мм. Верх рамы-полоса железа 30х3 мм длиной 72 см. Низ рамы длиной 80 см, концы ее (до 10 см) отогнуты. Посредине отогнутых концов сделано по отверстию диаметром 11 мм для оси трубы. Бока рамы сделаны из полосы длиной 42 см. Концы ее (по 6 см) отогнуты. Нa расстоянии 5 см от нижнего сгиба имеется отверстие диаметром 11 мм для оси трубы. Две скобы изготовляют из полос длиной 25 см, согнутых под прямым углом в четырех местах и прикрепленных к полозу у начала передней дуги. Две втулки для оси трубы делаются из полос длиной 28 см и имеют посредине отверстие диаметром 11 мм для оси трубы. Эти полосы сгибаются, вставляются в конец трубы, где закрепляются.
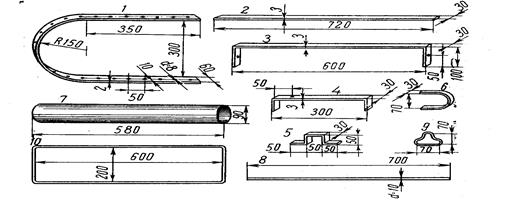
Рис. 35. Детали малой модели трала Расса (по Рассу, 1939).
Пояснения в тексте, условные обозначения те же, что и на рис. 34.
Труба представляет собой кусок железной газовой трубы диаметром 3,5″, длиной 58 см. Ось трубы делается из круглого железа диаметром 10 мм, длиной 70 см. Концы ее имеют винтовую резьбу для закрепления гайками. Две петли для уздечки изготовляются из куска круглого железа диаметром 10 мм идлиной 25 мм, как показано на рис. 36. Петли имеют внутренний прозор, равный 7 см. Рама для сети делается из 6-7-миллиметрового круглого железа длиной сторон 60 и 20 см. Бим представляет собой круглое бревно диаметром 7 см и длиной 72 см. Концы его четырехгранные и заточены (сторона 5 см) на 6 см от края. На четырехугольную железную раму надевается сетной мешок малой сети для лова икринок из газа № 15, имеющий в диаметре 50 см в длину и по боку 2 м.
Трал пускают на малом ходу судна. Вначале выбрасывают за борт конец сети трала со стаканом, тотчас же оттягиваемый Кодой назад, затем спускают весь трал. Ваер вытравляют на 2,5-4 глубины, пока трал не будет ползти по дну. Трал тянут обычно 5 мин от момента посадки на дно.
![]() Рингтрал (рис. 36) является основным орудием лова пелагических личинок и мальков. Он представляет собою коническую сеть из страмина, подвешенную на железном обруче. Диаметр обруча 2 м, длина сети 8 м. В собранном виде снабжается грузом, подвешенным у переднего конца оттяжек, и замыкателем. В последнее время стали пользоваться самописцем, отмечающим глубину, на которой идет трал.
Рингтрал (рис. 36) является основным орудием лова пелагических личинок и мальков. Он представляет собою коническую сеть из страмина, подвешенную на железном обруче. Диаметр обруча 2 м, длина сети 8 м. В собранном виде снабжается грузом, подвешенным у переднего конца оттяжек, и замыкателем. В последнее время стали пользоваться самописцем, отмечающим глубину, на которой идет трал.
Для экспедиции «Садко» В. Г. Богоровым был сделан рингтрал из шелкового газа, состоящий из следующих частей: обруча из круглого 15-миллиметрового железа диаметром 2 м, сетного корпуса и шелкового газа № 6 или 8, длиной 6 м (вдоль бока); четырех оттяжек, идущих вдоль всего трала и стакана.
Сетной конус надевается на обруч при помощи холщовой надставки. Поперечный полотняный (или холщовый) пояс вшит на расстоянии 4 м от обруча. От надставки к поясу вдоль бока конуса вшиты девять полотняных полос шириной по 2 см; от пояска до стакана-четыре полосы. Края полос сшиваются с полосами газа двойным швом. Рингтралом описанной конструкции можно производить горизонтальные поверхностные ловы или косые ловы от дна до поверхности на малом ходу. Для производства горизонтальных ловов в толще воды рингтрал надо прикрепить к ваеру какого-либо донного орудия (промысловый трал, трал Сигсби, драга), служащего в данном случае грузом, или же снабдить соответственным грузом, подвешенным перед входным отверстием.
Рис. 36. Рингтрал для
лова пелагических личинок и мальков (по Рассу, 1939).
Мальковый трал Петерсена (рис. 37) наряду с рингтралом может быть использован в качестве основного орудия лова пелагических мальков рыб. Он представляет собой сетной мешок с распорными досками, но без сетных крыльев, мешок трала имеет прямоугольный зев 230х130 см, причем бока его прикреплены к палкам длиной по 140 см, с окованными свинцом нижними концами. Вдоль боков, от зева к концу мешка, идут четыре веревочных растяжки. Конец мешка завязывается или снабжается специальным стаканом. Мешок изготовляется из тралина (19 ячей на 3 см), конгресс -канвы или мелкоячейной дели. Доски могут быть использованы те же, что и для малькового оттертрала. От концов боковых палок к углам досок идут по два линя длиной 6 м.
Пускают трал с наветренного борта на малом ходу, вытравляя тем больше ваера, чем более глубокие слои желательно обловить. Тянут трал 15-20 мин.
Мальковая лампара может быть рекомендована для лова пелагических и донно-пелагических сеголетков рыб. Пригоднa для мелких водоемов.
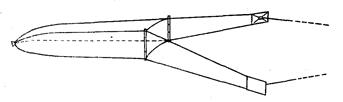
Рис. 37. Мальковый трал Петерсена ( по Рассу, 1939)
Высота лампары, применяющейся на Азовском море, равна 16 м, что позволяет в этом водоеме облавливать всю толщу воды-от поверхности до дна. Длина лампары (180 м по верхней подборе) позволяет небольшому судну делать при высыпке орудия замкнутый круг, длина которого равна длине лампары. Площадь облова такой лампары приблизительно равна 2500 м2.
Лампара применяется на моторных судах, имеющих длину корпуса от 15 до 24 м и обязательно свободную от построек корму, удобную для работы восьми человек при выборке.
Перед высыпкой лампары судно ориентируют так, чтобы дрейфом его относило от высыпаемой лампары. Приступая к лову, сбрасывают в воду бежной конец лампары длиной 20- 30 м с буем. Затем судно дает средний ход и описывает круг, возвращаясь к бую. Во время хода лампара, набранная на корме, постепенно выметывается в воду. К концу высыпки лампары буй и бежной конец выбирается на лине («сорочке») в носовой части судна, и судно подходит к крылу. Затем слегка стягивают нижнюю подбору (вручную или малым ходом судна), после чего оба крыла выбирают одновременно семь-восемь человек. Когда сак подходит к корме, рыбу сгоняют в один угол и поднимают с делью на борт (в случае небольшого улова). Весь процесс лова обычно длится 15-20 мин.
Для выбора наиболее уловистого и показательного места регулярных ловов необходимо предварительно произвести пробные ловы в разных местах. Ловы следует производить раз в 3 дня, причем, кроме дневного времени, весьма важны ловы в темное время, поздно вечером и ночью. Например, долгие поиски личинок мальков и сельди черноспинки (Caspialosa kessleri) у г. Саратова увенчались успехом лишь тогда, когда начали ловить молодь вечером. По этому поводу Б. И. Диксон (1905), занимавшийся исследованием сельди, писал: «Все дневные ловы (молоди сельди И. П.) до заката солнца были или абсолютно безрезультатными, или же давали одного-двух мальков, несмотря на многочисленные опускания сетки… С заходом солнца мальки внезапно появлялись в верхних слоях воды, и это появление обязательно совпадало с появлением лептодоры (Leptodora kindtii), поднимающейся с глубины на поверхность только при затенении плеса с заходом солнца».
У В. С. Танасийчук (1940) есть сообщение о лове молоди воблы. Молодь воблы ловили волокушей. Первое время среднюю часть волокуши делали из конгресс-канвы, а крылья из килечной дели. В дальнейшем вся волокуша была сделана из конгресс-канвы. Позднее эту волокушу заменили килечной из. 6-миллиметровой дели. Лов проводили обычно 2 раза в пятидневку на выбранном заранее месте, и каждый раз облавливали определенное расстояние (по берегу).
Глава VII
ОПРЕДЕЛЕНИЕ ВОЗРАСТА РЫБ
Значение исследований возраста рыб
Методика определения возраста рыб имеет обширнейшую литературу. Мы ограничимся изложением основ этой методики, наиболее доступных технических приемов определения возраста рыб и нахождения годовых приростов размера и веса и указанием соответствующих литературных источников, где научный работник найдет более подробные сведения по данному вопросу. Приводим также и некоторые критические замечания к работам по данному, весьма важному разделу ихтиологии.
При хозяйственном использовании объектов живой природы необходимо знать и возраст и годовые приросты (в весовых или линейных единицах) растения или животного.
Лесовод тщательно определяет возраст дерева по годичным слоям (кольцам), хорошо заметным на распиленном дереве. Но только по распилу корневого конца ствола можно узнать истинный возраст дерева. Пользуясь таким способом, лесники узнают и возраст отдельных деревьев, и средний возраст целой группы деревьев. Техника определения возраста деревьев хорошо разработана и упрощена. Слабо заметные годовые кольца дерева для надежного просчета смачивают красящими веществами.
Но важно знать не только количество годовых колец, но и их толщину. Одни кольца широкие, другие узкие, причем чередование широких и узких колец не имеет правильной закономерности: три кольца узких, затем два широких, далее одно узкое и три широких и т. д. Каждое широкое кольцо указывает, что во время образования этого кольца (в летнюю пору) дерево находилось в особо благоприятных условиях и дало большой прирост; узкие кольца говорят о слабых годовых приростах. Анализируя годовые кольца дерева, можно представить ход его жизни за прошлые годы, но для этого уже нужно уметь читать по этим кольцам жизнь дерева.
Лесоводы умеют определять возраст живых деревьев, не срубая их.
В животноводстве тоже без знания возраста животных невозможно вести правильные хозяйственные расчеты. Нужно знать и годовой прирост каждого животного. Этим знанием пользуются при выборе мясных, молочных и племенных животных. Всем известно, что по зубам определяют возраст лошадей, коров, свиней, верблюдов и других животных. Возраст млекопитающих можно достаточно точно определить и по срезам разных костей.
В. О. Клер (1927) приводит данные, свидетельствующие о том, что кости млекопитающих, как и кости рыб, имеют признаки, указывающие на прерывистость, периодичность их роста, и что по этим признакам можно определить возраст того животного, которому эти кости принадлежат.
Для рыбного хозяйства знание возраста и роста рыб имеет такое же значение, как и возраст и рост растений для лесного хозяйства и возраст и рост животных для животноводства. Нужно знать, в каком возрасте промысловая рыба становится способной к размножению, хорошо ли она растет, какие возрастные группы встречаются в улове и какие возрасты можно ожидать в будущих уловах.
Принцип определения возраста и роста рыб основан на свойстве чешуи и костей образовывать наслоения в виде чередующихся колец, поясов, плоскостей и склеритов-валиков (или гребешков). Каждому году жизни рыбы соответствует определенное кольцо на чешуе или на кости. Такие кольца по внешнему виду сходны с годовыми кольцами дерева и хорошо видимы даже невооруженным глазом. Не для всех рыб установлено точное время появления на теле чешуйного покрова, но у некоторых рыб первые чешуйки заметны через 3-4 десятка дней после выхода личинки из икры, у других значительно позднее. Чешуйка имеет вид тонкой прозрачной пластинки. Затем, как принято говорить, во второй год, хотя это не всегда соответствует календарному году, под первой пластинкой (под чешуйкой первого года) вырастает вторая, размером больше первой настолько, что ее периферический край выдается за такой же край первой пластинки. На третий год появляется третья и т. д. В конце концов, у взрослой рыбы, имеющей возраст, например, пять лет, каждая чешуя состоит из пяти пластинок (чешуек). Такое расслоение чешуи хорошо проследили на сазане В. В. Петров и Г. К. Петрушевский (1929). Таким образом, чешуя обнаруживает сходство с детской пирамидкой: вверху самый маленький кружок, внизу самый большой, сверху чешуи самая маленькая, она и самая старая пластинка, снизу-самая большая, она же самая молодая. Растет рыба в длину, растет и чешуя, а поскольку рыба в зависимости от условий среды и в зависимости, от состояния самой рыбы растет неравномерно, неравномерно растет и чешуя-то замедляясь, то ускоряясь. Эта неравномерность на чешуе выражается в виде различной толщины рубчиков, валиков, называемых склеритами (очевидно, от греческого слова склерос – жесткий, твердый), расположенными кругами или дужками. Ширина склеритов неодинаковая, неодинаковые и промежутки между склеритами. При быстром росте рыбы (летом) больше вырастает склеритов, расстояния между ними шире. При замедленном росте рыбы склериты сближены. Это наблюдается осенью. Считается, что зимой склериты вовсе не образуются (Чугунова, 1959). Полоса широких и полоса узких склеритов, образовавшихся в течение одного года, составляют годовое кольцо роста (рис. 38). Сколько на чешуе таких годовых колец, – столько рыбе и лет.
![]() Кроме годовых колец, на чешуе бывают дополнительные кольца, отображающие периоды изменений роста рыбы в течение года (см. ниже), зависящие от характера питания, температуры воды, времени нереста. Нужно различать еще мальковое кольцо, которое часто бывает заметно в зоне первого годового кольца. Форма чешуи у разных рыб разная, различны и рисунки чешуи, и распознавание колец роста требует хорошей наблюдательности и привычки исследователя.
Кроме годовых колец, на чешуе бывают дополнительные кольца, отображающие периоды изменений роста рыбы в течение года (см. ниже), зависящие от характера питания, температуры воды, времени нереста. Нужно различать еще мальковое кольцо, которое часто бывает заметно в зоне первого годового кольца. Форма чешуи у разных рыб разная, различны и рисунки чешуи, и распознавание колец роста требует хорошей наблюдательности и привычки исследователя.
Еще в XVII и XVIII веках некоторые естествоиспытатели видели сходство колец рыбьей чешуи с древесными кольцами и пытались по кольцам чешуи определить возраст рыб. Но только и XX веке изучение возраста и роста рыб получило строго научные основы.
Интерес к этому делу среди русских ихтиологов был пробужден кратким сообщением Н. А. Бородина в 1904 г. в журнале «Вестник рыбопромышленности». Практически определение возраста рыб в России впервые было осуществлено Е. К. Суворовым и И. Н. Арнольдом; первый в 1909-1910 гг. занимался определением возраста балтийских рыб (камбалы, салаки и кильки), второй в 1911 г. опубликовал статью с указанием методики определения возраста рыб (Арнольд, 1911). Эту статью по праву можно считать первым русским руководством по определению возраста рыб. Статья И. Н. Арнольда привлекла большое внимание и русских и иностранных ихтиологов. В. К. Солдатов в 1915 г. опубликовал замечательную книгу «Исследование осетровых Амура», где приводится определение возраста амурских осетровых по костям. Очень большое практическое значение имеет определение прироста рыбы за каждый год ее жизни, т. е. определение темпа роста рыбы. Возраст рыб и годовые приросты длины и веса рыбы мы определяем с достаточной точностью.
Если мы говорим, что в Каспийском море в весенних уловах воблы преобладают рыбы четырехлетнего возраста, что рыбы более старших возрастов встречаются реже, а восьмилетняя вобла является большой редкостью, то это еще не доказывает, что вобла живет только до восьми лет. В 70-е годы прошлого столетия много ,воблы ловилось в семи-восьмилетнем возрасте. В том и другом случае продолжительность жизни промысловой рыбы находилась в прямой зависимости от воздействия человека.
Долголетие, предельный возраст рыб легче установить в отношении рыб непромысловых или рыб в необлавливаемых водоемах, по и там приходится учитывать многие местные условия, которые мешают рыбам проявлять свой предельный возраст. О естественных предельных возрастах рыб мы знаем очень мало, но, несомненно, есть рыбы с весьма коротким жизненным циклом и есть долголетние рыбы. Например, горбуша живет обычно менее двух лет, ряпушка не дольше 10 лет, а белуги и калуги доживают даже до 75 и более лет. Большое значение, научное и хозяйственное, имеет вопрос о темпе роста рыб, и для решения его современная наука о рыбах действительно имеет надежные средства.
В чем же заключается значение точного определения возраста и роста рыб?
Сравнивая рыб одного возраста и одного и того же вида из разных водоемов, мы часто видим, что они имеют разную длину и разный вес. Например, трехлетние сиги одного и того же вида в одном озере имеют вес 900 г, а в другом водоеме- 400 г. Отсюда мы вправе заключить, что в первом озере сиги растут хорошо, в другом -плохо, или сиги в первом озере представляют форму быстрорастущих, а во втором-медленнорастущих. Нередко бывает и так, что сиги одного и того же возраста, взятые из одного и того же водоема, сильно разнятся по длине и по весу. Так, в Ладожском озере волховской сиг растет быстро, достигает крупных размеров, а сиг лудога крупным никогда не бывает. В таком случае приходится говорить о разных формах сигов: о волховском сиге следует говорить как о крупной форме, а о сиге лудоге как о мелкой.
В данном примере определение возраста приводит нас к двум важным результатам: а) кормность двух водоемов различна и б) в одном и том же водоеме имеются две формы рыб (сигов). Первый результат дает возможность сделать оценку -кормности водоема, а второй – оценку самой рыбы.
Ихтиологи, проводившие наблюдения над ростом ладожских и чудских сигов в озере Севане, нашли, что севанский сиг лудога превышает по росту лудогу из Ладожского озера. Ладожский рипус (крупная ряпушка), будучи вселен в приуральские озера, хорошо размножается там и имеет хороший темп роста.
Знание возраста рыб помогает определить скороспелые, быстрорастущие и медленно растущие виды рыб. В дальневосточных морях обитает горбуша, которая огромнейшими косяками входит во многие реки Камчатки, Сахалина, в реки Амурского лимана и Татарского пролива. Горбуша мельче других дальневосточных лососей (чавычи, кижуча, красной, кеты и симы), средний вес ее около 1 кг, но нерестится она всего лишь полуторагодовалой (Gilbert, 1912). Многочисленные материалы исследования возраста горбуши в реках Камчатки, Сахалина и в Амуре подтвердили это. Следовательно, изучением возраста горбуши была разгадана загадка о продолжительности жизни одной из главнейших промысловых и вместе с тем быстрорастущих рыб дальневосточных морей (к полутора годам горбуша становится уже совсем взрослой, половозрелой рыбой). Позднейшие исследования (Кагановский, 1949) показывают, что случается встретить горбуш и трехлетних, но эти особи, которые задержались в своем развитии, не нарушают приведенного выше общего заключения о раннем созревании горбуши.
В Амуре водится крупная, достигающая веса 1 т и более, калуга (Huso daurikus). Упомянутые великолепные исследования В. К. Солдатова (1915) показывали, что амурская калуга впервые мечет икру в возрасте 18-20 лет, имея вес почти в центнер. Значит калуга, в 10 лет имеющая значительный вес,- еще молодая, не достигшая половой зрелости рыба. Хозяйственники на основании этих данных должны считать калугу рыбой позднеспелой и, заботясь о поддержании калужьего промысла, не должны усиленно вылавливать калуг моложе 18 лет.
Стерлядь достигает половой зрелости на 3-4 году. А половая зрелость снетка наступает рано: однолетний снеток уже способен выметывать икру. Подобные сведения также необходимы для рыбного хозяйства, как для скотовода необходимо знание возраста разводимых им животных.
В зависимости от возраста рыб изменяются и некоторые признаки, отличающие один вид рыб от другого.
О значении изучения возраста рыб К. А. Киселевич (1927) говорит следующее: «беря из ходового косяка несколько сот или тысяч рыб и изучая чешую каждой из них, мы узнаем не только возраст каждой из них, но общее количество рыб 2,- 3-, 4-летнего и т. д. возраста. Для полноты картины всегда необходимо знать не только возраст рыбы, но и размеры ее, и за тем высчитать средние размеры каждого возраста. Беря из года в год в одних и тех же местах навсегда установленным способом по несколько сот или тысяч рыб и определяя их возраст и размеры, мы можем из года в год сопоставлять возрастной состав ловимой рыбы и средние размеры каждого возраста и проследить, насколько улучшаются или ухудшаются рыбные запасы». В другом месте он еще более выразительно говорит о значении исследования возраста рыб: «Изучая постоянно возрастной состав и темп роста рыб, мы можем судить о том, достаточно ли промысел использует природные запасы водоема и следует ли его (промысел) сокращать или расширять. На основе этих изысканий и строится рациональное рыбное хозяйство, т. е. наиболее полное использование рыбной продукции водоема». Короче говоря, без знания возраста рыб мы не можем судить о рыбных запасах того или другого водоема.
Определение возрастной группировки рыб необходимо там, где решается вопрос о недолове или перелове рыбы и вообще о запасах рыб. Средние пробы с преобладанием рыб старших возрастов (рыб переростков) .могут до известной степени служить подтверждением недоиспользования данных видов рыб. Уловы, состоящие из молоди рыб, указывают на перелов соответствующих пород рыб (если лов производится при одних и тех же условиях).
Зная быстроту роста рыбы, мы устанавливаем годовой (или даже месячный) прирост тела рыб, а это дает возможность определить возраст, в котором наиболее рентабельно, наиболее выгодно ловить данный вид рыбы. Известно, что старая рыба дает плохой прирост.
Каждому ихтиологу необходимо знать «Сборник статей по методике определения возраста и роста рыб» (1926), в котором дан ряд статей (переводных и оригинальных), имеющих высокую методическую ценность. Полезна статья из этого сборника Н. Л. Чугунова «Определение возраста и темпа роста рыб по костям». Автор, помимо теоретических обоснований, дает практические советы, описывает и разрабатывает метод, указанный В. О. Клером, расчисления роста рыб по костям. Кроме этого сборника, опубликованы книги и статьи, излагающие изучение роста. Особенно нужны работы Г. Н. Монастырского (1930 и др.), Н. И. Чугуновой ,(1959), Г. Г. Галкина (1958).
Определение возраста по чешуе
Годовые кольца на чешуе рыб обычно считают при увеличении 10-20-кратном, а иногда и более. В первом случае удобно пользоваться настольной лупой.
Не на всякой чешуйке хорошо различимы кольца роста. Поэтому надо отобрать хорошую чешую, толково ее сберечь и приготовить для подробного просмотра. При этом требуется выполнять следующие правила сбора.
Записать название рыбы, место и время сбора, размер и вес рыбы. Указать всю длину рыбы (ab), длину тела до конца хвостовой выемки (ас), длину тела без хвостового плавника (ad) и длину туловища или тушки (od). Эти промеры обозначены на рис. 39. Вес рыбы показать в граммах или килограммах.

Рис. 39. Измерения при исследованиях роста и возраста рыб.
Пунктиром показаны места, откуда следует брать чешую (по Правдину, 1939).
Очень существенно определить пол и степень зрелости половых продуктов.
На основании таких определений можно судить, на каком году самцы и самки становятся половозрелыми, а также о сроках их нереста.
У большинства рыб чешую для определения возраста берут с середины бока рыбы (в границах пунктирных овалов, рис. 39), повыше или пониже боковой линии. Возраст бесчешуйных рыб определяют по костям, а чешую с рыб, не имеющих боковой линии, берут с середины бока рыбы.
С каждой рыбы берут по 5-10 чешуек и кладут в конвертики или в особую книжечку (размером 5-10 см) из писчей бумаги.
Собранные чешуи хранят в сухом месте. При определении возраста чешуйки промывают в разведенном нашатырном спирте или в простой воде (сырой) и очищают мягкой щеткой (или между пальцами) от покрывающей их слизи.
Возраст определяют обычно по передней части чешуи.
Прежде чем приступить к определению возраста рекомендуется тщательно ознакомиться с многочисленными формами чешуи разных возрастов рыб, изображенными в Атласе Г. Г. Галкина (1958).
В статье А. И. Рабинерсона (1927) говорится о структуре чешуи норвежской сельди (рис. 40): «Вся поверхность чешуи изборождена параллельными рядами тонких черточек слегка изгибающихся по краям; эти линии производят впечатление: как будто это дуги окружностей, описанных очень большими радиусами, во всяком случае во много раз превышающими размеры чешуи.

Такие образования, носящие названия склеритов, у сельди располагаются довольно однообразным рядом и никаких указаний на возраст особи не дают». Кроме того, на чешуе резко бросаются в глаза четыре концентрических темных кольца. Эти четыре кольца дуги, не что иное, как границы годовых колец чешуи.
Первое (от центра) кольцо соответствует полному первому году жизни рыбы, второе – второму году и т. д. Годовые кольца на чешуе сельди выражены на передней части чешуи. На задней части они незаметны. На рис. 41 изображена чешуя трехлетней сельди. Годовые кольца ограничены белыми (светлыми) линиями.
Для большей наглядности на рис. 42 дана схема расположения склеритов амурской горбуши, взятой в 1928 г. В центре чешуи небольшое колечко а1 со склеритами, не сильно разобщенными. Далее идет кольцо с редко расположенными склеритами а2. Оба кольца (а1+а2) мы считаем за одно кольцо А, первое светлое кольцо. За этим кольцом-пояс сильно сближенных склеритов, темное кольцо Б. Наконец, последний пояс редко сидящих склеритов, светлый пояс В, окаймляющий край чешуи.
Таким образом, горбуша, взятая в 1928 г., имела возраст (я об этом упоминал выше) около полутора лет, т. е. являлась поколением, вышедшим из икры, отложенной осенью 1926 г. Осенью 1926 г. горбуша вошла в реку и отложила там икру. Из икры к весне 1927 г. вывелись мальки горбуши, которые некоторое время пожили в реке (колечко a1) и затем скатились в море (колечко a2). Между колечками a1и а2, т. е. между жизнью малька в речной воде и в морской, есть маленький поясок сближенных склеритов. Это сближение склеритов можно подметить, если внимательно отсчитывать склериты первого кольца А от центра чешуи к ее верхнему и нижнему краям (сближены 7, 8 и 9-й склериты). Очевидно, при переходе из пресной воды в соленую рост склеритов замедлился (пока рыба привыкала к новой среде обитания). При таком толковании образование всего первого кольца (А) относится к периоду март-сентябрь 1927 г., образование второго кольца (Б) – к октябрю-декабрю 1927 г. и январю-марту 1928 г., а последнее (незаконченное) кольцо (В) – к апрелю-июлю 1928 г. Следовательно, возраст взятой горбуши, пришедшей в Амур для нереста, мы устанавливаем в полтора года.
Рис.42. Схематический рисунок чешуи горбуши. Возраст полтора года (по Правдину, 1939)
На рис. 43 дан фотографический снимок чешуи горбуши. Итак, при счете годовых колец за годовое кольцо принимают два кольца: одно-светлое с крупными склеритами и одно (соседнее) темное с мелкими склеритами.
У атлантических лососей Salmo salar и S. trutta хорошо различимы речные и морские кольца роста, а так как эти лососи могут нерестовать несколько раз (обычно не более 3 – 4) то у них на чешуе отличают еще так называемое нерестовое кольцо ![]() или нерестовую марку. Перед изучением возраста лососей по чешуе нужно просмотреть микрофотографические снимки их чешуи; такие рисунки во множестве имеются и в советских и в иностранных изданиях (рис. 44).
или нерестовую марку. Перед изучением возраста лососей по чешуе нужно просмотреть микрофотографические снимки их чешуи; такие рисунки во множестве имеются и в советских и в иностранных изданиях (рис. 44).
На рис. 45 показан сильно увеличенный отрезок задней части чешуи сига. На отрезке чешуи семь годовых колец. Первое годовое кольцо расположено в центре чешуи и состоит из нескольких ( 10-12) кругов полосок, достаточно разделенных между собой. Круги это и есть те склериты, о которых шла речь выше. Первое кольцо отделено от второго годового кольца
Рис. 44. Чешуя лосося после первого нереста (по Суворову из Чугуновой, 1952).
(здесь говориться о годовых кольцах, состоящих из ряда склеритных колечек, а не о кольцах отдельных склеритов) темной полосой, по обе стороны видно по несколько рядов весьма сближенных, узко расставленных склеритов. Такими же темными Полосами отделены и следующие кольца: второе от третьего, третье от четвертого и т. д. На рисунке легко подметить, что годовые кольца не одинаково широки: первые (считаем от центра) три годовых кольца шире, чем последующие четыре, причем, последнее кольцо, седьмое- самое узкое. Следовательно, в первые три года чешуя росла быстро, а начиная с четвертого

года рост чешуи, очевидно, стал замедляться. Кольцо седьмого года, вероятно, не закончено, но все же определенно можно сказать, что ширина седьмого кольца будет меньше любого из трех первых годовых колец. Подмеченная неравномерность годовых колец объясняется тем, что сиг быстро рос в первые три года; на четвертом году он, очевидно, нерестился, вследствие чего рост его был замедлен; медленно по сравнению с первыми тремя годами протекает рост и других, более старших возрастов (5, 6 и 7 лет). К этому вопросу мы вернемся, когда будем говорить о темпе роста рыб.
Рис. 46 заимствован из книги финского исследователя Т. Н. Ярви (Jarvi, 1928). В этой книге подобных рисунков (еще более отчетливо изображенных) много, и на каждом из них можно сосчитать годовые кольца. В работах советских авторов (особенно последних лет) также имеется много рисунков чешуи сигов и других рыб.
Рассмотрим еще чешую рыбы из семейства карповых. На рис. 46 показана чешуя трехлетней воблы, на чешуе три годовых кольца (Чугунова, 1959). Здесь также ясно выражен характер склеритов и отчетливо видно их расположение.
Мы уже знаем, что на чешуе сельди склериты расположены в виде очень отлогих дуг, а на чешуе лососей и сигов-в виде довольно правильных овалов или даже кругов. Кругами они выражаются и на чешуе карпа.
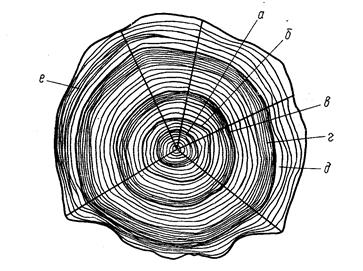
Рис. 46. Схема чешуи трехлетней рыбы (по Чугуновой, 1952).
На чешуе выражены, помимо годовых колец (а, в и д), добавочные кольца (б, г и е).
Светлые кольца считаются летними кольцами, определяющими летний рост рыбы, а кольца темные-зоны замедленного роста, часто называемые зимними кольцами. Следовательно, в каждом годовом кольце нужно различать эти две части. Задним краем годового кольца считают задний край кольца сближенных склеритов. Раньше кольцо сближенных склеритов принимали за зимнее кольцо, теперь этот термин более соответствует осеннему кольцу (Чугунова, 1959), но и он не менее условен, чем термин зимнее кольцо.
Посмотрите еще раз на приведенные здесь рисунки чешуи. На чешуе норвежской сельди зимние концентрические кольца не совпадают ни с расположением, ни с размером склеритов. На чешуе горбуши зимние кольца трудно отделить от летних; зимними кольцами мы здесь считаем группу сближенных склеритов. Примерно такой же рисунок имеют зимние кольца чешуи и многих других (но не всех) рыб семейства лососевых. На чешуе карповых зимние кольца, совпадая с кольцами склеритов, вместе с тем резко отграничены от летних колец и так же, как и у лососевых рыб, состоят из ряда сближенных узких склеритов. Нужно заметить, что название «зимнее кольцо» можно употреблять не всегда, .потому что рост многих рыб зимой приостанавливается.
От каждой исследуемой рыбы необходимо просмотреть нeсколько (во всяком случае не менее 5-7, а в сомнительных случаях и значительно больше) чешуек. Чешуя молодых камбал очень удобна для определения возраста (рис. 47).
Рис. 47. Чешуя четырехлетней камбалы (по Правдину, 1939).
Помимо неясностей чешуи, возникающих вследствие повреждений чешуи или уклонений от нормального вида (бывают чешуйки, где никаких колец подметить невозможно), очень часто (у некоторых рыб вполне закономерно) можно подметить упомянутые выше нерестовые кольца.
На примере чешуи горбуши было указано, что в первом летнем кольце горбуши имеется маленькое дополнительное колечко, появление которого мы объяснили замедлением роста чешуи (и всей молодой рыбы) вследствие временного неблагоприятного состояния рыбы при переходе ее из речной воды в морскую. Но в жизни рыб могут быть и другие причины, также способствующие временному замедлению роста. К таким причинам относится и нерест рыбы. Во время созревания половых продуктов и в процессе их откладывания, которому порой предшествуют весьма длительные путешествия к местам нереста, сопровождающиеся у многих рыб голоданием, рыба худеет, и ее рост и рост чешуи замедляются. Склериты последней в период нереста рыбы (от момента подготовки к нересту) узки и дают темные кольца, которые нельзя считать за годовые. Такие добавочные кольца затрудняют и иногда сильно путают счет годовых колец на чешуе. Неопытный наблюдатель может ошибиться и преувеличить число лет. Но если их понять, то добавочные кольца позволяют объяснить многие явления жизни рыб.
Нерестовые кольца или нерестовые марки подмечены на чешуе многих рыб. Из советских ученых подробно изучили нерестовые марки на чешуе сельдей Д. Ф. Замахаев (1940) и Н. И. Чугунова (1940 а.). Д. Ф. Замахаев отметил, что на чешуе нескольких видов каспийских сельдей откладывается нерестовая марка, характеризующаяся такими признаками: 1) присутствие кольца слома, не параллельно краям чешуи, 2) резкое пересечение кольцом слома рядов стрий (тонких неконцентрических складок на передней части чешуи; задний край чешуи стрий не имеет); 3) отсутствие стрий в проксимальной части зоны нового прироста; 4) нарушение направления новых стрий на некоторых участках нового прироста и 5) резкое утончение зоны нового прироста. Нерестовые марки у сельдей наиболее заметны на чешуе, расположенной под спинным плавником по середине тела.
Еще лучше выражены нерестовые кольца на чешуе лососей. По некоторым кольцам делают заключения и о годе первого нереста рыбы и о повторных нерестах.
Таким образом, по чешуе определяется не только возраст рыбы, но и другие жизненные явления и справедливо предложение Н. И. Чугуновой употреблять термин «чтение чешуи».
Закладка годового кольца не всегда начинается ранней весной (в зависимости от температуры). В этом отношении представляет интерес работа Н. С. Соловьевой (1938) по изучению времени закладки зимних колец и начала прироста на чешуе сельди в различных районах Баренцева моря. Автор пришел к выводу, что «годовое кольцо на чешуе мурманской сельди начинает закладываться в разгар гидрологического лета (август). Таким образом, связь прекращения роста рыбы с понижением температуры в данном случае отсутствует, очевидно главное влияние на закладку зимнего кольца оказывает прекращение питания, что наступает вследствие высокой степени упитанности сельди. Рост чешуи, а следовательно, и самой сельди начинается в мае, когда она, израсходовав запасы жира за зиму, получает громадное количество пищи и усиленно откармливается».
Распознавание зимних колец-очень трудная работа: исследователь всегда сталкивается с многочисленными переходными формами этих колец, но принцип, положенный в основу подобных различий, представляет несомненный интерес. Аналогичные исследования зимних колец сельди проведены П. А. Мурашкинцевой (1938).
У каждого вида рыб годовые и добавочные кольца имеют свои особенности, которые могут быть выяснены путем просмотра массового количества чешуи.
Н. И. Чугунова (1959) дает указания по различению колец на чешуе воблы.
Годовые кольца замкнуты, идут параллельно окружности чешуи и образуются они на границе тесно расположенных склеритов осенне-зимнего роста и раздвинутых склеритов весенне-летнего роста. Нерестовые кольца обычно заметны по разрыву склеритов и неправильному их расположению: обрывки склеритов идут в разных направлениях. На задней части чешуи нередко образуется утолщенный темный склерит, который обычно разбит на подкововидные отрезки.
Добавочные кольца образуются под влиянием различных факторов среды (резкое изменение физических условий среды, питания и др.). Добавочные кольца очень похожи, но менее отчетливо выражены, чем годовые и бывают нескольких типов (мальковые, кольца случайной задержки, или наоборот, усиления роста и т. п.). Мальковые или покатные кольца образуются недалеко от центра чешуи (внутри первого годового кольца). Вероятно, возникает такое кольцо при переходе малька из реки в море. Границы малькового кольца менее резко выражены, чем границы первого годового кольца.
Мальковое кольцо не для всех рыб обязательно, но установление его необходимо для того, чтобы точно определить кольцо первого года жизни рыбы: нередко мальковое кольцо принимают за годовое. Н. И. Чугунова (1959) дает такой ответ: «Когда впервые рассматривают чешую какого-нибудь вида рыбы, возраст и рост которой неизвестен, приходится сопоставлять длину сеголетков, собранных в водоеме осенью, или годовиков, собранных ранней весной, с расчисленными по чешуе длинами рыбы в возрасте одного года. Таким путем удается установить разницу между годовым и мальковым кольцами». Этот способ может быть принят весьма условно. И сам автор признает, что первое годовое кольцо должно быть распознаваемо по его структуре, а не по соответственной расчисленной длине рыбы, так как если длину рыбы в возрасте первого года находить по обратному расчислению длин крупных рыб, то она, как правило, будет меньше нормальной.
При выращивании сеголетков сазана в волжской пойме нами (мною и Ф. Э. Карантонисом) было подмечено, что на чешуе сазана 4-5-месячного возраста было несколько хорошо отграниченных колец, которые, если бы неизвестен был истинный возраст сазана, следовало бы признать за годовые кольца.
Ф. Э. Карантонис (1945) дал исчерпывающие объяснения этих дополнительных колец. Чешуя сазана, прожившего всего одно лето, имела 6 колец и каждое кольцо соответствовало особенностям условий жизни молоди. Первое кольцо постларвального периода (возраст около 10 суток после выхода из икры, длина тела 10 мм). Второе кольцо полойного периода, когда рыбка росла интенсивно, пользуясь просторами залитых водой лугов; за 12 суток полойной жизни рыбка получает прирост в 42 мм. Третье кольцо-склериты сильно сближены- у молоди, сазана, оставшейся в высыхающих озерках, где условия существования для молоди сазана были исключительно плохи. Четвертое кольцо возникло у рыбок, отсаженных нами в пруд, где они получали обильный корм. За 39 дней пребывания в пруду мальки выросли на 107,5 мм. Пятое кольцо указывает на новое угнетение роста вследствие ухудшения питания (рыбки были лишены искусственного подкармливания). Шестое кольцо-кольцо обычной осенней депрессии роста, свойственной всем рыбам нашего климата.
Если видимость годовых колец на чешуе неясная, то одним из. способов улучшения видимости их считается так называемая дифференцированная окраска чешуи, которая предложена П. В. Тремповичем (1932). Чешую, завернутую в марлю, выдерживают в течение 17-20 ч в растворе (37,5%) сернокислого железа. Перед исследованием чешую хорошо промывают обычной (водопроводной) водой, обсушивают фильтровальной бумагой и переносят в каплю раствора (3%) таннина.
От действия сернокислого железа и таннина чешуя чернеет. Годичные кольца становятся более заметными. Однако не у всех рыб такое окрашивание чешуи дает нужные результаты. Например, у уклей (Милинский, 1946), у которой зимние кольца очень узки (состоят всего из двух рядов узких склеритов), такое окрашивание не улучшает видимости годовых колец.
Возраст рыб обозначается или римскими или арабскими цифрами без плюса или с плюсом (8,8+). Первая восьмерка показывает, что рыба имеет полных 8 лет (например, окунь, взятый в мае-июне, имеет полные года), вторая восьмерка (8+) указывает, что рыбе более восьми лет, но менее девяти (например, окунь, взятый в декабре). Возраст лососей обозначают иначе: впереди ставится число лет, проведенных лососем в реке, а затем указывается число лет, проведенных в море (или в озере). Например, 3+1 означает, что в реке лосось (молодь) прожил три года, затем один год он жил в море. Иногда такую формулу обозначают иначе: 3+1+SM+1, т. е. лосось в реке провел три года, затем один год в море, потом нерестился в реке (знак SM) и снова один год провел в море. Может быть, и такая запись 5з, 64 и т. п. Строчные числа указывают на общее число лет лосося, подстрочные – на число лет проведенных 1 реке. Встречаются в ихтиологической литературе и другие Приемы обозначения лет жизни лососей.
Определение возраста по костят и отолитам
На многих костях рыб, как и на чешуе, правильно чередуются полоски. Одни из этих полосок кажутся (даже при просмотре без увеличительного стекла) светлыми, другие -темными. Светлые полоски -широкие, темные-узкие, т. е. наблюдается картина, повторяющая рисунок чешуи.
![]() В результате изучения возраста по костям было установлено, что лучшим материалом для этого служат кости плоские, похожие на пластинки. Такими костями у рыб являются четыре кости жаберной крышки – предкрышечная, крышечная, покрышечная и межкрышечная, челюстные кости, окаймляющие рот, кости так называемого плечевого пояса, отделяющие жаберную щель рыб от туловища, а также плоские Кости черепа (рис. 48).
В результате изучения возраста по костям было установлено, что лучшим материалом для этого служат кости плоские, похожие на пластинки. Такими костями у рыб являются четыре кости жаберной крышки – предкрышечная, крышечная, покрышечная и межкрышечная, челюстные кости, окаймляющие рот, кости так называемого плечевого пояса, отделяющие жаберную щель рыб от туловища, а также плоские Кости черепа (рис. 48).
Способ определения возраста рыб по костям быстро вошел в практику исследователей рыб: приводились и приводятся все новые и новые данные, свидетельствующие о том, что на костях каждой рыбы имеется строго определенное количество полосок, совпадающее с годовыми кольцами чешуи, и что эти полоски точно определяют число лет жизни рыб. Помимо указанных выше костей, при определении возраста рыб берутся позвонки и косточки из слухового аппарата рыб, известные под названием отолитов, или слуховых косточек, а также жесткие лучи плавников.
И. Н. Арнольд (1911) привел данные по определению возраста белуги, осетра, севрюги, щуки, окуня, судака, налима, сазана, леща, плотвы, воблы, линя, язя, сырти или рыбца, и сигов. Для счета лет осетровых рыб И. Н. Арнольд рекомендует брать кости плечевого пояса и жаберной крышки, причем последнюю необходимо утончить, отшлифовать; возраст щук хорошо определять и по отшлифованным позвонкам, у окуня годовые кольца следует считать по крышечной когти и по кости верхней челюсти, задний край которой у окуня заканчивается значительным расширением; у налима-по позвонкам и отолитам; у сазана-по жаберной крышке и позвонкам; у леща и плотвы-по костям плечевого пояса; у рыбца-по позвонкам; у сигов-по жаберной крышке и позвонкам.
Указаниями И. Н. Арнольда в основном пользуются до сих пор все, кто занимается изучением возраста рыб по их плоским костям. А. Г. Смирнов (1929) отметил, что годовые полосы на костях жаберных крышек аральской шемаи выражаются далеко не ясно. Поэтому автор провел специальное исследование жаберных крышек шемаи. Он окрашивал кости гематоксилином, метиленблау, пикрокармином и борным кармином и подвергал действию 25% аммиака, 5-10% едкого натра и едкого кали, бензола, бензина, серного эфира, этилового спирта и глицерина. Только обработка глицерином дала заметную пользу. Очищенные жаберные кости в течение 10-15 мин выдерживались в глицерине, затем в нем же нагревались до 290°С (т. е. до кипения). В кипящем глицерине кость из прозрачной становится молочно-белой, и на этом фоне годовые кольца начинают отчетливо выделяться. При дальнейшем кипячении объект желтеет и в связи с этим картина годовых наслоений несколько затемняется.
В. К. Солдатов (1915), выполнивший труднейшую работу по определению возраста очень большого количества осетровых рыб Амура, пишет: «При обработке костей мы поступали так: осторожно вырезав те кости, которые нам требовались, из свежей рыбы, мы не надолго, чтобы только легко отделились мышцы и другие мягкие части, опускали их в кипящую воду ключом; отделив от костей все ненужные части, промыв кости водой и протерев щеточкой, мы их обычно высушивали и сохраняли в сухом виде до того момента, когда их нужно было просматривать. Обычно хорошо вываренные и предварительно очищенные кости, возможно, было сразу же рассматривать, так как слоистость на них выступала весьма рельефно: для других костей требовалась их дальнейшая обработка – спиртом различной концентрации и бензином или эфиром для удаления из них влаги и жира. Самый просмотр производился так: кости слегка смачивались спиртом и рассматривались на свет или же на темном фоне, смотря по толщине и большей или меньшей четкости в чередовании слоев».
На основании определения возраста осетровых Амура В. К. Солдатов пришел к интересным, имеющим огромное хозяйственное значение, выводам. «Калуга становится взрослой рыбой, способной к размножению, только через 17 лет по выходе из икры, достигнув к этому времени не менее 5 пудов (около центнера) весом и около 230 сантиметров полной длины или 165 сантиметров промысловой, а амурский осетр делается способным к воспроизведению себе подобных на 9-10-м году по выходе из икры, достигая к этому времени в среднем около 14 фунтов (около 6 кг) весом при абсолютной длине в среднем около 108-116 сантиметров полной и около 73,4-78,8 сантиметров промысловой длины».

![]() Те же выводы дали В. К. Солдатову основание решительно Говорить о том, что существовавший в то время на Амуре промысел добывал калуг и осетров преимущественно малолетних, т. е. не достигших половой зрелости. Из просмотренных В. К. Солдатовым более 2000 калуг неполовозрелых оказалось около 91%, а из 6 тысяч (с лишним) осетров неполовозрелых было 94%. Нерациональность такого промысла очевидна и явилась причиной резкого сокращения запасов осетровых Амура. Для восстановления запасов советской Властью был проведен полный запрет на лов амурских осетровых.
Те же выводы дали В. К. Солдатову основание решительно Говорить о том, что существовавший в то время на Амуре промысел добывал калуг и осетров преимущественно малолетних, т. е. не достигших половой зрелости. Из просмотренных В. К. Солдатовым более 2000 калуг неполовозрелых оказалось около 91%, а из 6 тысяч (с лишним) осетров неполовозрелых было 94%. Нерациональность такого промысла очевидна и явилась причиной резкого сокращения запасов осетровых Амура. Для восстановления запасов советской Властью был проведен полный запрет на лов амурских осетровых.
А. Н. Пробатов (1936) на основании изучения возраста и роста осетровых Амура по костям (по поперечным шлифам первого луча грудного плавника) пришел к выводу, что в Амуре имеются обособленные биологические группы калуг и осетров.
На рис. 49-53 показаны очищенные кости рыб с заметными на них годовыми приростами.
![]() Широко распространен способ определения возраста рыб по отолитам. У рыб нет наружного и среднего уха, т. е. нет ни ушной раковины, ни слухового отверстия, ни барабанной перепонки, а есть только так называемое внутреннее ухо со слуховым нервом. Внутри слухового аппарата лежат и отолиты, имеющие у различных рыб различную форму.
Широко распространен способ определения возраста рыб по отолитам. У рыб нет наружного и среднего уха, т. е. нет ни ушной раковины, ни слухового отверстия, ни барабанной перепонки, а есть только так называемое внутреннее ухо со слуховым нервом. Внутри слухового аппарата лежат и отолиты, имеющие у различных рыб различную форму.
Рис. 52. Кость плечевого пояса двенадцатигодовалого
леща (по Арнольду, 1911),
Найти отолиты в головной части рыбы без навыка не так легко, но приобрести нужный навык для каждого вполне доступно. Возьмите две-три сушеных головы корюшки или ерша, у которых косточки головы мелки и легко рассыпаются. Среди размельченных костей вы быстро усмотрите два крупных белых зернышка продолговатой формы; одна сторона этих зернышек выпуклая, другая-вдавленная, к наружным краям зернышек идут бороздки. Эти зернышки и есть отолиты. Присмотревшись к отолитам, можно легко отыскать их место и расположение среди головных костей и на свежих рыбах.

Рис. 53. Межкрышечная кость двадцатипятилетней камбалы (по Арнольду, 1911).
На отолитах ясно выражены годовые кольца, по которым и определяют возраст рыб (рис. 54). Тщательное изучение отолитов камбалы немецким ученым Иммерманом показало, что отолит камбалы состоит из студенисто-волокнистого вещества, в котором содержатся кристаллы углекислой извести. Волокна отолитов имеют способность спаиваться в концентрические пластинки, причем весной и летом вырастают белые кольца, осенью- темные. Белое кольцо вместе с темным считают за одно годовое кольцо. Хорошо различаются кольца на отолитах молодых рыб, хуже-на отолитах рыб старых.
Многие современные работы, относящиеся к определению возраста рыб, содержат немало новых указаний, как и какими костями следует пользоваться при изучении возраста рыб. Но обычно просматривают не одну какую-либо кость, а берут разные кости и чешую, чем и достигается контроль правильности определения. Поэтому отбирать и просматривать приходится и отолиты.
![]() П. Ф. Федоров (1931) подверг химической обработке отолиты беломорской корюшки и свои выводы описал так: «Прежде всего, отолит помещается в 25% аммиак (известный под названием нашатырного спирта), способствующий его обезжириванию. В аммиаке отолит выдерживается от 30 мин до 24 ч, но в большинстве случаев 4-5 ч. После такой обработки отолит промывается в горячей воде и затем рассматривается под лупой в капле глицерина». Однако такая операция не всегда дает хорошие результаты: были случаи, когда отолит выдерживался в аммиаке более суток и годовые кольца его все же не становится более ясными.
П. Ф. Федоров (1931) подверг химической обработке отолиты беломорской корюшки и свои выводы описал так: «Прежде всего, отолит помещается в 25% аммиак (известный под названием нашатырного спирта), способствующий его обезжириванию. В аммиаке отолит выдерживается от 30 мин до 24 ч, но в большинстве случаев 4-5 ч. После такой обработки отолит промывается в горячей воде и затем рассматривается под лупой в капле глицерина». Однако такая операция не всегда дает хорошие результаты: были случаи, когда отолит выдерживался в аммиаке более суток и годовые кольца его все же не становится более ясными.
Рис. 54. Отолиты (сверху вниз):
отшлифованный отолит пятигодовалого
налима; отолит шестигодовалого налима;
отолит шестигодовалой камбалы.
Такие отолиты после выдерживания в аммиаке и промывки горячей водой клались на 3-5 мин в кипящий раствор поваренной соли (на 6 г соли бралось 100 см3 воды) и снова промывались в горячей воде. Видимость годовых колец от этого улучшалась.
Если описанный способ обработки не придавал годовым кольцам отолита требуемой отчетливости, то П. Ф. Федоров изготовлял шлифы. По линии, проходящей перпендикулярно продольной оси отолита, производился разрез лобзиком через центр (продольный распил отолита) или просто отолит стачивали напильником, а затем тщательно шлифовали на оселке. После такой шлифовки отолит заливали в канифоль на предметном стекле так, чтобы отшлифованная плоскость была обращена к стеклу. Когда канифоль затвердевала, напильником спиливали вторую часть отолита до тех пор, пока оставшаяся пластинка его на стекле становилась близка к просвечиванию. Затем пластинку отолита шлифовали на оселке до полной просвечиваемости. Когда шлиф был готов, на предметное стекло вводили каплю ксилола, растворявшую канифоль. Затем объект заливали канадским бальзамом и покрывали покровным стеклом.
А. Я. Базикалова, Т. Н. Каллиникова, В. С. Михин и Д. Н. Талиев (1937) определяли возраст байкальских бычков по шлифам отолитов. Отолиты шлифовались с обеих сторон на куске прессованной пемзы. Хорошие результаты дает шлифовка на белой мелкозернистой искусственной пемзе хорошего качества; крупнозернистая грубая пемза непригодна для шлифовки, так как на ней шлиф, не будучи еще достаточно тонким, легко ломается.
Шлифы отолитов бычков просматривались в глицерине на предметном стекле при слабом увеличении микроскопа и при проходящем свете.
Отолиты используются как материал для определения не только возраста, но и роста трески. Д. Ф. Замахаев (1941) описал методику расчисления роста трески по отолитам, признав, что при определении возраста и роста старых рыб отолиты имеют большое преимущество перед чешуей: лучше видны годовые кольца и быстрей можно вести обработку (за 6 ч можно приготовить 40 препаратов чешуи, а отолитов за то же время – около 200 штук). Автор так описывает этот метод.
Отолит предварительно покрывался черным лаком, разламывался руками и отшлифовывался на точильном камне. Обращалось особенное внимание на то, чтобы слом по возможности приходился по середине отолита, на ровность шлифа и на перпендикулярность плоскости шлифа к продольной оси отолита.
Кольца на отолите измеряли окуляр-микрометром под бинокулярном в отраженном свете.
Годичные кольца на шлифе отолита можно измерять в двух направлениях – по короткой и длинной осям.
В направлении короткой оси измерения производились на вогнутой стороне отолита, по длинной – на суженной его части. В первом случае точка, от которой производились промеры, часто не совпадала с центром первого годового кольца, во втором случае измерения производились от этого центра. Нужно отметить, что центр первого годового кольца не всегда совпадает с центром отолита. Трудно сделать такой препарат, относительно которого была бы полная уверенность, что центр начального роста отолита попал в плоскость шлифа.
Измерение годичных колец по длинной оси шлифа менее удобно ввиду их расплывчатости и большого количества трещин, которые образуются преимущественно в этом направлении при разломе и шлифовке отолита. Наоборот, измерения по короткой оси не представляют никакого затруднения.
Оказалось, что разница в расчислениях роста по обоим измерениям очень незначительна. Д. Ф. Замахаев пришел выводу, что отолиты вполне пригодны для определения роста трески длиной от 30 до 100 ом.
В. О. Клер (1916) предложил совершенно новый метод определения возраста рыб-метод, который в настоящее время стал весьма распространенным. Он установил, что «возраст рыб удобнее читать не на плоских цельных костях, как это обычно принято, а на костях, имеющих компактную структуру, что в свою очередь возможно только при изучении костей посредством разрезов шлифов».
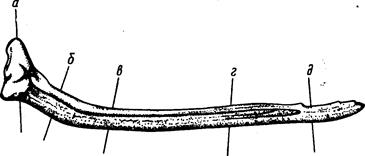
Рис. 55. Первый луч грудного плавника стерляди (по Клеру, 1916). Распилы луча по линиям а, б, и в дали правильное определение возраста.
В этих словах выражена вся сущность клеровского метода определения возраста рыб. Этот метод дает возможность брать для определения возраста осетровых такие кости, как первый луч грудного плавника или косточки, лежащие у верхней лопасти хвостового плавника, так называемые фулькры. Названные кости легко вырезать, и рыба (товар) от такой операции нисколько не портится.
На рис. 55 изображен первый луч грудного плавника крупной стерляди. Буквы а, б, в, г и д указывают места, где были сделаны поперечные распилы. Распилы по линиям а, б и в дали правильное определение возраста стерляди; на распилах г и д возраст был меньше, потому что распил не захватил старых, первых годичных слоев. На рис. 56 изображен шлиф пластинки из того же луча; на шлифе хорошо заметны 10 годовых колец.
В. О. Клер (1927 а), занимаясь исследованием структуры костей разных животных, много сделал и для методики определения возраста рыб.
Если у осетровых рыб хорошо видны годовые кольца на шлифе первого луча грудного плавника, то у нельмы (из лососевых рыб) возраст легко определить на шлифе верхнечелюстной кости. На рис. 57 представлены верхнечелюстные кости нельмы (прямой линией обозначено место распила). На рис. 58 ясно видны годовые кольца на шлифе отрезка верхнечелюстной кости нельмы.
![]() Ценные указания сделал В. О. Клер и по технике изготовления шлифов. Лучшим шлифовальным порошком он считает размельченную пемзу.
Ценные указания сделал В. О. Клер и по технике изготовления шлифов. Лучшим шлифовальным порошком он считает размельченную пемзу.
Рис. 56. Отшлифованная пластинка первого грудного плавника десятилетней стерляди (по Клеру, 1916). Цифрами указаны годовые кольца.
Шлифовать костяные пластинки следует в вазелиновом или другом жидком минеральном масле. Для окончательной отделки шлифа при наклейке на стекло В. О. Клер рекомендует употреблять канадский бальзам. «Для этого берется канадский бальзам, высушенный до такой твердости, что при комнатной температуре при надавливании ногтем на нем остается лишь небольшой след. Для приклейки стекло нагревается настолько, чтобы при накладывании к нему куска бальзама последний плавился. Операция производится на медной пластинке, нагреваемой спиртовой лампочкой».
![]()
Рис. 57. Верхнечелюстные кости нельмы (по Клеру, 1916). Прямая линия, идущая через шейки костей, обозначает место, где следует делать распил.
Д. Н. Талиев (1931) впервые применил способ определения возраста трески (рыбы с мягкими лучами) по первому лучу спинного плавника. Автор брал луч вместе с суставной головкой (у трески он состоит из двух валиков). Шлиф изготовлялся так. Начиная от суставной головки, луч шлифовали на полубархатном напильнике, держа его за верхнюю часть; когда на напильнике вышлифована суставная головка и почти весь перехват, заканчивали шлифовку на матовом стекле в наждаке или на мелкозернистом бруске, наблюдая при этом, чтобы луч был всегда перпендикулярен шлифующей поверхности.
На отшлифованной поверхности через увеличительное стекло нетрудно подсчитать годовые кольца. Для получения двустороннего шлифа Д. Н. Талиев прикреплял при помощи расплавленной на спиртовой лампе канифоли луч отшлифованной поверхностью; по остывании канифоли, чтобы не шлифовать весь луч, его обламывали внизу у канифоли. Для изготовления одного шлифа требуется 3- 10 мин. Возраст на таких шлифах определяют под микроскопом при малых увеличениях. Препарат плавникового луча показан на рис. 59.
Рис. 58. Поперечный шлиф распила верхнечелюстной кости нельмы (по Клеру, 1916). Ясно видны годовые кольца.
Возраст сомов по первому лучу грудного плавника определяется тем же способом, как и возраст осетровых рыб. «От каждого луча у головки, в том месте, где оканчивается углубление, выпиливалась лобзиком пластинка толщиной до 1 мм и отшлифовывалась на мелком напильнике», – так описывает А. Н. Пробатов (1929) подготовку лучей сома для определения его возраста.
Рис. 59. Шлифы первого и второго лучей первого спинного плавника четырехгодовалой трески ( по Талиеву, 1931).
Рис. 60 изображает поперечный разрез луча грудного плавника восьмилетнего сига, рис. 61-пятилетнего жереха.
В методику определения возраста осетровых рыб по костям внесено много существенного Н. Л. Чугуновым (1926), предложившим простой способ изготовления нужных для определения возраста пластинок лучей осетровых рыб. Изготовление поперечного среза луча производится обыкновенным лобзиком, в котором строго параллельно закрепляются две тонких пилки, разделенные в зажимах лобзика тонкой медной пластинкой. При помощи установленных таким образом пилок без труда выпиливается соответствующей толщины срез, который при надлежащей установке пилок не требует и последующего шлифования.
![]()
![]()
Рис. 60. Поперечный шлиф луча грудного плавника восьмилетнего сигапо Пробатову, 1929).
Рис. 61. Поперечный шлиф луча плавника восьмилетнего сига грудного плавника жереха (по Пробатову, 1929). Жереху 5 лет.
Для большей ясности в просчете годовых плоскостей пластинки лучей следует перед просмотром погружать в ксилол.
Толщину среза следует делать около 0,5 мм. Выпиливать срез Н. Л. Чугунов советует вблизи сочленовой головки луча и не далее 1-1,5 см от нее. Просчет годовых колец ведется под лупой или под микроскопом. Для выпиливания наиболее пригодны самые тонкие пилки по металлу.
Вместо лобзика с двумя параллельно вставленными пилками стали употреблять специальный прибор. Прибор для изготовления срезов лучей осетровых описан Н. И. Чугуновой (1959).
Он представляет собой площадку, на которой укреплены две поставленные параллельно (на расстоянии 0,5 см) дисковидные мелкозубчатые пилки из инструментальной стали. Эти пилки приводятся в движение ручным способом или электричеством. Луч плавника кладется в зажим, установленный на столике перед пилками, который при помощи пружины, находящейся под столиком, подвигает луч к пилкам (рис. 62).
Срез делается у самой головки, чтобы сохранить на срезе первый год. Первый срез делается не далее 1 см от переднего края головки. Возраст на таких срезах (отшлифованных в том же приборе) определяют при увеличении в 20-25 раз. Срезы |ля просветления смачивают толуолом или ксилолом.
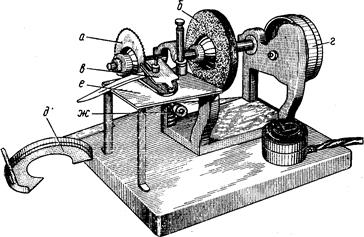
Рис. 62. Прибор для изготовления срезов лучей осетровых, вид спереди (по Чугунову, 1926):
a-дисковая пилка; б-шлифовальный круг; в-зажимы; г – передача от электромотора: д – защитный кожух; е – столик; ж – винт.
Л. П. Астанин (1947) описал новый способ приготовления срезов недекальцинированных костей. Кости размягчаются под влиянием варки в воде или под действием пара. Потом такие кости легко режутся бритвой.
П. А. Дрягин (1936), встретившись с трудностями определения возраста сазана из р. Чу, пользовался шлифами срезов третьего жесткого луча спинного плавника.
В. В. Петров (1927) применил более удобный способ сбора лучей осетровых. Он обертывал вырезанный луч полосками бумаги, на которой записывал измерения и пол рыбы, и затем клал кости на солнце без всякого вываривания. Кости высыхали, и бумага приклеивалась. В таком виде кости завертывались в общий пакет и укладывались в ящик для отправки.
Д. А. Бельчук (1938) занимался определением возраста одноперого терпуга. Для этой цели были взяты кости жаберной крышки operculum и suboperculum, чешуя, кость плечевого пояса-cleithrum, позвонки и отолиты. Лучшую видимость и более точное количество лет имела чешуя, взятая под грудными плавниками и со средней части тела. Отолиты оказались слишком хрупкими и желаемых результатов не дали. На suboperculuni годичные наслоения видимы лучше, чем на operculum, но более удобной для определения возраста оказалась кость cleithrum. Просчет лет по позвонкам очень затруднителен. Автор приходит к выводу, что лучшими элементами для определения возраста терпуга является чешуя и cleithrum.
Работами Доно-Кубанской научно-исследовательской рыбохозяйственной станции внесены изменения в методику определения возраста рыб по спилам плавников (Бойко, 1951). Доказана возможность получения поперечных разрезов плавниковых лучей разных рыб (и не только осетровых, сома и тресковых). Спилы готовились из целого плавника, а не только из первого простого луча). Е. Г. Бойко так описывает методику изготовления спилов плавниковых лучей.
Отделяется и засушивается весь плавник или несколько лучей плавника. При высушивании лучи плотно скрепляются друг с другом плавниковыми перепонками и при распиливании не крошатся. Плавники с тонкими лучами (у мелких рыб) для уплотнения заливаются в целлулоид. Поперечный спил делается при помощи лобзика с тонкой пилкой. Вместо шлифовки спилов применяется просветление их трансформаторным и репейным маслами или ксилолом. Толщина спила 0,4-0,5 мм. Годовые кольца видны и на простых и на ветвистых лучах. Хорошей видимостью обладают плавниковые спилы жереха, леща, судака, трески, карпа, карася, чехони, тарани, густеры, рыбца, вырезуба, язя, кеты, семги, иваси, каспийского пузанка, волжской и донской сельдей, кефали и лобана. Годовые кольца на лучах не были обнаружены у пеламиды. Автор пишет, что за 7-8 ч можно приготовить 200-250 спилов или залить в целлулоид 200-300 плавников, или просмотреть до 300-500 спилов.
Плавниковые лучи от мелких рыб, прежде чем изготовлять из них спилы, можно утолщать, опуская несколько раз в раствор фотопленки в ацетоне. Пленку предварительно промывают в теплой воде для удаления эмульсии.
Методика использования плавниковых лучей для определения возраста рыб более пригодна, чем по чешуе и костям, и может производиться быстро и без большой подготовки. Однако неясен вопрос о возможности применения таких спилов для расчисления темпа роста рыб.
Нельзя не отметить попытку С. М. Кагановской (1933) определить возраст акулы Squalus acanthias по колючкам спинных плавников; на этих колючках имеются полосы, соответствующие годовым кольцам.
Описанные способы определения возраста рыб по чешуе и по костям – самые надежные, но они ни в какой мере не являются окончательными: исследовательская мысль не должна успокаиваться, а должна проверять, критиковать эти способы, существенное из них углублять, а устаревшее или не отвечающее современному состоянию ихтиологической науки отбрасывать и заменять новым, более обоснованным.
![]() Однако до сих пор встречается необходимость пользоваться методами более простыми. К числу таких методов можно отнести так называемые петерсеновские кривые, употребляющиеся для быстрой хотя и далеко не точной) группировки улова рыбы по ее (возрастному составу (рис. 63).
Однако до сих пор встречается необходимость пользоваться методами более простыми. К числу таких методов можно отнести так называемые петерсеновские кривые, употребляющиеся для быстрой хотя и далеко не точной) группировки улова рыбы по ее (возрастному составу (рис. 63).
Предложенный норвежским исследователем Петерсеном прием впервые вошел в практику научно-промысловых рыбных исследований более 50 лет назад и состоит в следующем.
Рис. 63. Петерсеновская кривая.
Из улова берется порция (по возможности большая) рыб. Каждая рыба измеряется, затем показатели длин группируются в классы, например, через 1-2-5-7 и т. д. см, и подсчитывается, сколько в каком классе рыб. Может получиться, например, такой ряд:
Длина 0 10 20 30 40 50 60 70 80 90 100 110 120 130 140 150
тела, см
Число 10 | 53 | 100 |120 | 70| 5 | 0 | 0 | 5 | 50 | 80 | 70 | 90 | 20 | 5
особей
Рассматривая приведенный ряд, можно подметить, что во взятой пробе нет рыб, длина которых была бы от 60 до 80 см. В пробе есть рыбы мелкие и крупные, т. е. взятая проба включает рыб молодых и старых (имеется в виду, что проба относится к одной и той же систематической группе). Полученный ряд можно более наглядно представить графически (см. рис. 63). График показывает то же, что и ряд, но простота и наглядность на графике несравненно больше. Хорошо видно, что улов рыбы делится на две группы: первая имеет длину тела от 10 до 60 см, вторая-от 80 до 150 см. Первая группа-молодые рыбы, вторая – рыбы взрослые. Такие же ряды можно составлять и по весу рыб.
Группируя этим способом рыб по длине их тела, мы приближаемся к выводам о возрасте рыб, но только приближаемся, потому что точный возраст взятых рыб остается, не определен; не определен также и темп роста этих рыб.
Глава VIII
ОПРЕДЕЛЕНИЕ ТЕМПА РОСТА РЫБ
Метод и приборы для обратного расчисления темпа роста
Рост рыбы не прекращается с наступлением половой зрелости. Но в молодом возрасте рыба обычно растет быстрее, чем в старшем. Известно, что в периоды созревания половых продуктов, когда много веществ в организме расходуется на выработку этих продуктов, в период миграции и нереста, когда многие рыбы перестают питаться, рост рыбы замедляется и даже вовсе приостанавливается.
Несмотря на то, что методы изучения темпа роста рыб продолжают разрабатываться многочисленными авторами, уже теперь от определения линейных приростов рыбы (приростов в длину) замечается переход к определению весовых приростов, т. е. ихтиологи в этом вопросе приближаются к животноводам, которые ценность домашних животных определяют главным образом весовыми показателями.
Норвежский исследователь Эйнар Леа (Lea, 1910), изучавший возраст и темп роста норвежской сельди, пришел к заключению, что рост чешуи пропорционален росту рыбы, т. е. годовой прирост чешуи так же относится к длине чешуи, как годовой прирост всего тела (длины тела) к общей длине тела. Иначе говоря, если найдено, что чешуя за определенный год выросла в какой-либо избранной нами части на 1/10 всей длины взятой чешуи, то нужно полагать, что за тот же год прирост всей рыбы в длину составлял тоже ‘/10 длины тела. Если мы определим соотношение годовых приростов чешуи за каждый год, то тем самым сможем определить какая длина тела рыбы соответствовала каждому (любому) прожитому ею году.
Зная длину рыбы, длину чешуи (обычно исчисления ведутся не по всей чешуе, а по части ее, лежащей от центра до наружного или внутреннего края) и ширину годовых колец, можно определить длину тела рыбы за все предыдущие годы ее жизни (рис. 64). Этот способ определения годовых приростов рыбы называется методом обратного расчисления роста рыбы. Вычисления ведут по особой формуле, имеющей несколько сражений. Самое простое обозначение этой формулы
![]()
где L – длина рыбы;
С – длина чешуи (от центра до края в той части, где определяются годовые кольца);
Сх – длина чешуи за первый год (от центра чешуи и включая первое годовое кольцо); этим же выражением обозначается величина чешуи за два, три и т. д. года;
lx– длина рыбы за первый, второй, третий и т. д. годы.
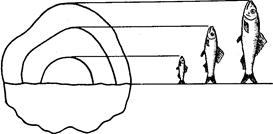
Рис. 64. Соотношение между ростом чешуи и длиной рыбы (по Правдину, 1939).
Следует соблюдать однообразное буквенное обозначение ветчин приведенной формулы. Н. Л. Чугунов (1926) говорит: «Вычисленные длины рыбы для первого, второго и т. д. годов принято обозначать l1, l2, l3 и т. д., а буквой L-полную длину исследуемой рыбы. Кроме того, для характеристики величины прироста рыбы по отдельным годам – так называемого темпа роста – вычисляют эти приросты, обозначаемые t, путем вычитания величины рыбы предшествующего года из величины последующего: прирост первого года t1=l2; t2=l2–l1; t3= l3–l2 и т. д.». Замечание это весьма важно, и его следует выполнять при работах по изучению темпа роста рыб. Приведенную выше формулу поясним на примере.
Возьмем четырехлетнюю рыбу, имеющую длину тела (L) 30 см и расчислим темп ее роста. Измерим чешую в том месте, где считали годовые кольца – например, от центра чешуи к ее краю по задней, свободной части чешуи. Наметим для 4 лет кольцо шириной 5 мм, для 3 лет – шириной 4 мм, для 2 лет – 3 мм и для 1 года – 2 мм. Какой размер имела рыба, когда ей было 3 года?
Решаем формулу, заменяя буквы нашими цифрами.
![]()
Длина тела рыбы в 3 года ее жизни была 24 см; в 2 года
![]()
за первый год-
![]()
Следовательно, длины тела рыбы, полученные путем обратного расчисления (в см), таковы: за 1 год l1– 12, за 2 года l2– 18, за 3 года l3-24. За 4 года l4–30 см по непосредственному (эмпирическому промеру).
Так как l4 обозначает длину тела рыбы по нашему промеру, то эту величину (полную длину тела) следует обозначать буквой L.
Найденные цифры показывают еще и то, что рыба в длину за каждый год (кроме первого года) прибывала по 6 см, что ее темп роста за второй, третий и четвертый годы шел равномерно, давая одинаковые годовые приросты, именно t2– 6 см., t3, – 6 см и t4 – 6 см.
Целый ряд ихтиологов проверяли и пытались уточнить метод Э. Леа. Многие не соглашаются с тем, что рост чешуи рыбы прямо пропорционален росту тела рыбы в длину. Г. Н. Монастырский, например, проанализировав зависимость между длиной тела рыбы и размерами чешуи, пришел к выводу, что прямой пропорциональности между ростом чешуи и ростом рыбы в длину нет, и предложил определять темп роста рыб по чешуе при помощи логарифмических шкал, а также ввел в практику ихтиологов прибор для механического вычисления темпа роста рыб.
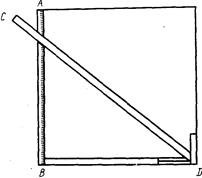
Рис. 65. Доска Эйнара Леа для обратного расчисления темпа роста рыб (по Правдину, 1939). Рисунок схематический (на левом крае доски должны быть миллиметровые деления).
Описание этого прибора станет более понятным, если сначала мы дадим описание той расчетной доски, какую сконструировал Э. Леа. Доска Э. Леа (рис. 65) несколько измененная, достаточно понятно описана М. И. Тихим (1917). «На квадратной доске, составленной во избежание прогиба из нескольких досок, по левому краю расположена латунная шкала шириной 14-20 мм с точным делением на миллиметры.
Величина шкалы, а равно и доски, зависит от наибольшей величины объекта, рост которого изучают. Вдоль нижнего края привинчена такая же линейка толщиной 2-3 мм, но без деления. Перпендикулярно к последней по нижнему ребру доски прикреплена латунная полоса, часть которой выступает над плоскостью доски на 5 мм. По этому выступу передвигаются особого рода «салазки». Салазки, закрепленные винтом, представляют вершину угла, вокруг которого вращается гипотенуза треугольника, в данном случае – массивная линейка. 0бе ее стороны срезаны для того, чтобы облегчить чтение цифр по шкале. Конец линейки, обращенный к салазкам, скошен, заострен и делается из твердого металла. Он упирается в угол, образуемый салазками, и выступает нижней, расположенной на ребре доски, полосой».
Для расчетов роста рыбы по доске необходимо проделать следующую работу. Чешуя рыбы, именно ширина ее годовых колец, проектируется через рисовальный аппарат на прямоугольник из бумаги или тонкого картона (наиболее удобный размер прямоугольника-4 см х 10 см), причем левый нижний край этой полоски бумаги совмещают с центром чешуи, а на полоске делают отметки (карандашом) границ каждого годового кольца. Детали работы легко уяснить при пользовании доской.
Допустим, что чешуя взята у рыбы, длина которой 450 мм. Возраст этой рыбы 5-й год (4 полных года и 5-й незаконченный). Полоску с нанесенными отметками устанавливаем на доске, помещая ее нижнюю кромку в угол, образованный выступом нижнего ребра доски и нижней полоской. Гипотенузную линейку ставим одним (острым) концом в угол салазок, а другой конец, именно его нижнюю линию, совмещаем с делением миллиметровой шкалы, с 450 мм. Полоску бумаги с отметками подводим под гипотенузу таким образом, чтобы первая (сверху) отметка (отметка 5-го года) совпала с нижней линией гипотенузы. Затем, не передвигая картона с отметками, гипотенузу перемещаем до совпадения со второй отметкой, соответствующей четырем годам рыбы. Левый край гипотенузы покажет на миллиметровой шкале размер тела рыбы за то же время, т. е. за время, когда рыбе было 4 года. Точно таким же приемом мы узнаем длину тела рыбы за 3, 2 и 1 год.
Определение возраста с успехом производится с помощью микроскопа. Так, Г. X. Шапошникова (1935) определение возраста молоди семги вела при 80-кратном увеличении. Для обратного расчисления чешую измеряли окуляр-микрометром с помощью передвижного столика микроскопа. «Данные по измерению переднего края чешуи и длины рыбы (по Смитту) подставлялись в формулу
![]()
где Ln искомая возрастная длина, Vn–расстояние между зимними кольцами, V-длина чешуи в делениях окуляр-микрометра от центра до края и L – измеренная длина рыбы». Техническое выполнение расчетов можно вести или по расчетной (логарифмической) линейке или на счетной машине: длину рыбы делят на всю длину чешуи в делениях окуляр-микрометра и полученный коэффициент помножается на длину чешуи от центра до зимних колец.
Если нужно определить годовые приросты рыбы, то поступают обычным порядком, т. е. вычитают длину тела рыбы за предшествующий год из длины тела за последующий.
Помимо чешуи для обратного расчисления берут и кости рыб (шлифы костей): плечевого пояса, жаберной крышки и лучей. Сущность расчетов по костям та же, что и расчетов по чешуе, но пользование костями требует несколько иных приемов, описание которых имеется в упоминавшейся выше статье ,Н. Л. Чугунова (1926).
Шлифы грудного луча осетровых признаются пригодными не только для определения возраста этих рыб, но и для обратного расчисления, однако, как говорит К. Г. Дойников (1936), для обратного расчисления пригодны не все шлифы лучей, а только те, на которых плоскости роста хорошо видны. При расчислении темпа роста азовской севрюги К. Г. Дойников пользовался боковыми ветвями (лопастями) луча на шлифах, так как на этих ветвях зимние кольца наиболее четко выражены. Годовые кольца отдельных шлифов при помощи рисовального аппарата наносились на карточки, затем расчислялись на доске Эйнара Леа.
Методика определения линейного роста взрослого (промыслового) налима по отолитам разрабатывалась и впервые описана А. В. Лукиным (1935). Отолит зарисовывался под лупой с помощью рисовального аппарата Аббе. Затем с точностью до 1 мм измерялись продольная и поперечная оси отолита, а также расстояние от центра отолита до наружной точки периферии по той половине поперечной оси, где годовые кольца выражены наиболее заметно. Расчисление роста велось методом Монастырского (методом логарифмических шкал), и автор пришел к выводу, что в данном случае этот метод «дает достаточно точные результаты»,
Г. Н. Монастырский (1924) сконструировал расчетную доску, более известную под названием доски Монастырского (рис.66), Главное отличие доски Монастырского от доски Э. Леа в том, что расчет ведется не по отметкам на карточках (где карандашные линии имеют некоторую ширину и тем самым увеличивают ошибку в точности вычислений), а непосредственно по проекции чешуи.
Доска имеет форму прямоугольного треугольника. Слева к ней прикреплена такая же миллиметровая шкала, как и на доске Э. Леа. Гипотенузная линейка здесь заменена натянутой черной тонкой нитью FO. К доске прикреплен столик, на который ставится микроскоп или лупа с рисовальным аппаратом. Последующие вычисления ведутся по принципу доски Э. Леа. Этот прием показан на рис. 67.
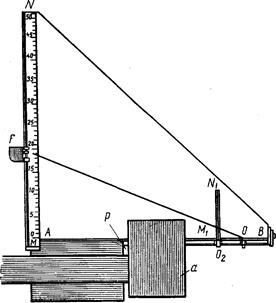
Рис. 66. Расчетная доска Монастырского – Э. Леа (по Монастырскому, 1924):
NMB-треугольная доска; NM-металлическая измерительная шкала; F- движок; OF – гипотенузная нить, соединяющая движок F с муфтой О, скользящей по металлическому валику, параллельному АВ; АВ-прямая линия, прочерченная на белой плотной бумаге, наклеенной на доску; О2-вторая муфта, к которой прикреплена белая узкая металлическая линеечка длиною в 10 см, на линейке черная тонкая линия N1M1, строго перпендикулярная линии АВ; а – столик движущийся, на который устанавливается лупа или микроскоп.
Проекция чешуи своей средней линией должна совпадать с линией N1M1, которая прочерчена на металлической пластинке. Центр чешуи совпадает с точкой пересечения линии N1M1 с АВ, а край чешуи должен касаться натянутой линии FO.
Дальнейшая разработка метода обратного расчисления привела Монастырского к мысли пользоваться логарифмическими шкалами, и он предложил новый прибор для механического вычисления линейного роста рыб по чешуе. Схема прибора изображена на рис. 68.
Сущность прибора и способ пользования им описываем словами Г. Н. Монастырского (1930): «Прибор имеет форму квадратной доски со сторонами 30 см. Параллельно нижнему краю, на расстоянии 5 см, неподвижно прикрепляется логарифмическая шкала АВ. Продолжение этой шкалы посредством шарнира соединяется с вращающейся линейкой АС, которая на свободном конце имеет продольный вырез для зажимного приспособления. Это приспособление помещается в вертикальном пазе bс. Длина линейки АС равна 37 см.
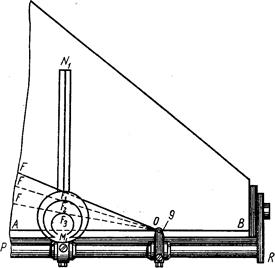
Рис. 67. Применение доски Монастырского (по Монастырскому, 1924).
Параллельно шкале АВ на расстоянии 1 см проходит паз с закраинами, в котором ходит движок de, имеющий перпендикулярный паз для другой логарифмической шкалы ED. Благодаря такому приспособлению шкалу ED можно удерживать в вертикальном положении, а также можно передвигать параллельно и вертикально по отношению к шкале АВ.
Деления на шкале АВ отвечают значениям измерений чешуи, а деления на шкале ED отвечают измерениям рыбы. Линейка АС соответствует прямой, проходящей возле эмпирических точек на логарифмической сетке.
Пользоваться прибором очень просто. Сначала устанавливается линейка АС, которая располагается под тем же углом относительно шкалы АВ, что и прямая на логарифмической сетке по отношению к оси абсцисс, т. е к горизонтальной оси. Затем шкала ED подводится к делению на шкале АВ, соответствующему выбранному измерению чешуи, и передвигается вверх и вниз до пересечения отметки, отвечающей длине рыбы, с нижним обрезком линейки. После этого шкала ED ставится против делений, соответствующих значениям расстояний от центра чешуи до зимних колец, и каждый раз замечается, на каком делении шкала пересекается с линией Л С».

Рис. 68. Схема прибора Монастырского для вычисления длины рыбы по чешуе (по Монастырскому, 1930):
АВ и D-логарифмические шкалы; AC-вращающаяся линейка; de-движок.
При отсутствии логарифмического счетного прибора Г. П. Монастырского можно пользоваться весьма несложным приспособлением, предложенным Г, П. Померанцевым (1949), позволяющим производить обратные расчисления длины и веса рыб по логарифмическим шкалам. Описание этого приспособления и техники работы с ним приведено в работе Н И Чугуновой (1959).
Г. П. Померанцев рекомендует расчислять длины и веса не по отдельным особям, а по средним величинам, полученным для каждой возрастной группы рыб, что значительно экономит время и не снижает точности результата.
Прибор Г. Н. Монастырского в последних работах русских ихтиологов находит применение, но все же до сих пор более распространен метод Эйнара Леа, не превзойденный по своей простоте и для многих рыб дающий хорошие результаты.
В. В. Петров (1935), определяя темп роста иссык-кульского сазана, пишет: «Темп линейного роста изучался методом обратного расчисления, предложенным Hjort и Е. Lea; другие, часто более точные, но и более кропотливые методы (Шерифа, Монастырского, Ли) нами не применялись, ибо, как показано было рядом работ (Борзенко, 1924; Пожалуева, 1928; Петров, 1931, 1932), влияние возраста изучаемых рыб на точность обратного расчисления для сазана чаще совсем не сказывается, а если в редких случаях и имеется намек на существование «феномена Розы Ли», то этот «феномен» всегда выражен слабо и мало влияет на степень точности расчисления».
Ю. Н. Редкозубов (1951) ввел значительные упрощения в технику обратного расчисления темпа роста рыб по чешуе и сконструировал прибор, представляющий собой модификацию доски Леа – Монастырского (рис. 69).
В основу предложенного Редкозубовым прибора положен принцип подобия треугольников и признания прямой пропорциональной зависимости между длиной тела рыбы и размерами чешуи (на таких же принципах построена и доска Эйнара Леа и Г. Н. Монастырского).
Перпендикулярно шкале, сделанной из полоски миллиметровой бумаги и наклеенной на доске через нулевое деление шкалы, натянута струна между двумя планками, на внутренних верхних углах которых выбран фалец. Концы планок, удаленные от шкалы, связаны замыкающей дощечкой. Обращенные к шкале концы планок врезаются в доску так, чтобы их верхние плоскости совпадали (заподлицо). Один конец струны закрепляется наглухо за штифт, вбитый в замыкающую дощечку. Штифт вбивается несколько выше линии оси, а для того, чтобы струна проходила точно по линии оси, к замыкающей дощечке с внутренней стороны прикрепляется направляющая планка оси (углы планки надо слегка закруглить, чтобы она не резала нить). Ходовой конец струны, пройдя через нулевое Деление шкалы, пропускается через отверстие.
Струна натягивается при помощи натягивающего приспособления, устроенного по принципу скрипичного колка. С внутренней стороны отверстия, для того чтобы струна не резала дерево, врезается металлическая планочка. По наружному краю доски сверху и снизу выбираются две направляющих канавки, по которым ходят направляющие штифты движка с укрепленным на нем барабанчиком. На барабанчик наматывается нить. Основой движка служит деревянная колодочка, к которой привинчиваются
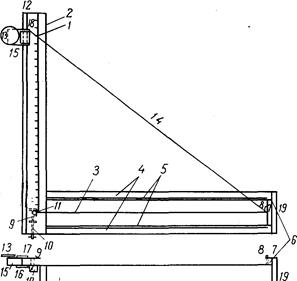
Рис. 69. Расчетная доска Редкозубова (по Редкозубову, 1951):
1-шкала; 2-доска; 3-струна; 4-планки; 5-фалец; 6-замыкающая дощечка; 7-штифт; 8-направляющая планка; 9-отверстие; 10-натягивающее приспособление; 11– металлическая планочка; 12 – направляющие канавки; 13-барабанчик; 14-нить; 15-деревянная колодочка; 16 и 17-нижняя и верхняя планки; 18-штифт; 19-ножки; 20-опорная доска.
Барабанчик легко можно сделать, спаяв его из толстой жести или склеив из бумаги. В последнем случае крышки барабанчика делаются из жесткого картона. Внутри барабанчика вставляется пружина от карманных часов, которая, вращая его, держит нить все время в натянутом состоянии. При конструировании прибора наибольшее затруднение было встречено при подборе нити, которая должна быть достаточно эластичной и прочной. После многочисленных опытов выбор остановился на тонкой капроновой нитке.
Барабанчик Нить Верхняя планка
![]()
Направляющие штифты
Колодочка Нижняя планка
![]()
Нить
Направляющие
штифты
Барабанчик Колодочка Верхняя планка
Рис. 70. Детали расчетной доски Редкозубова (по Редкозубову, 1951).
Собранный прибор ставится да три ножки, две из которых, лежащие под началом и концом струны, связаны опорной доской.
Для работы с прибором требуется подвижной стеклянный столик, изготовляемый из обычного предметного стекла. Вдоль его длинной стороны посередине прочерчиваются две параллельных линии, перпендикулярных к короткой стороне. Столик вставляется между планками в фалец таким образом, чтобы струна проходила поверх стекла, а линии, прочерченные на столике, были перпендикулярны к ней. В случае затруднения с прочерчиванием линий на стекле их можно начертить на куске прозрачной фотопленки, которая затем приклеивается к стеклу. Поверхность стеклянного столика должна приходиться на одном уровне с поверхностью планок и доски со шкалой. Это достигается или выборкой на соответствующую глубину фальца или фалец выбирается .заведомо слишком глубоким, а к столику вдоль его коротких сторон приклеиваются соответствующей высоты ножки из плотного картона или дерева.
Для работы с прибором пригодна любая препаровальная лупа. Лупу ставят на опорную доску, причем высота ножек должна быть такой, чтобы столик Лупы подходил под планки (для удобства столик можно снять). Расстояние от наружной кромки планки до струны делается таким, чтобы струна приходилась против центра зеркала лупы.
При работе с крупной чешуей, например сазана, семги, леща, сига и т. п., можно обойтись без приготовления специального препарата. Чешую надо только отмыть от слизи и грязи и высушить в расправленном состоянии. При работе с мелкой нежной чешуей (например, сельди) изготовляют специальные препараты. Для этого берут неиспользованную фотографическую пленку и, не помещая в проявитель, в темноте обрабатывают фиксажной солью. Этим достигается прозрачность покрывающего пленку желатинового слоя. Отрезав нужный кусочек обработанной подобным образом пленки, на ее поверхность, покрытую желатином, капают каплю, воды и |на размягченный этим желатин наклеивают отмытую чешуйку. Необходимо строго следить за тем, чтобы чешуйки легли на желатин гладкой стороной, а ребристой – вверх. Чешуйку придавливают мягкой тряпочкой и дают препарату высохнуть. Изготовленные подобным образом препараты долговечны, портативны и обходятся дешевле, чем препараты, смонтированные между двумя стеклами.
Изготовленный препарат или отдельную чешуйку накладывают на подвижной стеклянный столик прибора, заправляя его под струну. Струна должна быть так натянута, чтобы могла крепко придерживать препарат на столике. Центр чешуи совмещают с точкой перекреста струны с одной из линий, прочерченных на столике. Дальнейшая работа производится так же, как и на первой модели доски Монастырского: движок передвигают вдоль шкалы до пересечения нити с делением, соответствующим длине рыбы. Столик вместе с препаратом передвигают по разам до совмещения края чешуи с нитью. Необходимо только следить, чтобы при установке центр чешуи был совмещен с перекрестком струны и линии на столике. Лупу устанавливают так, чтобы луч, отраженный от зеркала лупы, проходил через чешую. Передвигая движок вдоль шкалы до совмещения нити с соответствующим годовым кольцом, получаем искомую длину рыбы.
Для проверки достаточности натяжения нити рекомендуется приготовить тест-препарат. На продолговатом куске прозрачной фотопленки прочерчивается длинная ось, пересеченная произвольным числом поперечных штрихов. Наложив тест-препарат на столик и взяв произвольный исходный отсчет на шкале, совмещают последний поперечный штрих с нитью и производят расчисления, как по чешуе. Производя расчисления при движении движка сверху вниз, доводят движок до нулевого деления шкалы и производят вторично отсчет при движении движка снизу вверх. Идентичность отсчетов будет служить показателем достаточного натяжения нити. Для лучшей установки при работе полезно слегка подергивать нить пальцем.
![]() При изготовлении прибора размеры будут лимитированы имеющейся в распоряжении лупой. Тем не менее, можно рекомендовать следующие основные размеры: длина шкалы длина оси, перпендикулярной шкале (струны) 550-600 мм; диаметр барабанчика примерно 40 мм; материалом могут служить сосновые доски толщиной 10-20 мм; струну следует брать самую тонкую. 500 мм;
При изготовлении прибора размеры будут лимитированы имеющейся в распоряжении лупой. Тем не менее, можно рекомендовать следующие основные размеры: длина шкалы длина оси, перпендикулярной шкале (струны) 550-600 мм; диаметр барабанчика примерно 40 мм; материалом могут служить сосновые доски толщиной 10-20 мм; струну следует брать самую тонкую. 500 мм;
Рис. 71. Схема углового масштаба Алеева (по Чугуновой, 1952):
С – центр, СА – радиусы, ВВ – вертикальные линии, по которым ведутся отсчеты.
Необходимое требование при работе с предлагаемой доской – употребление длиннофокусной лупы. Недостаток употребления длиннофокусной лупы и некоторая громоздкость доски не имеют существенного значения в лабораторных условиях.
Удобен, хотя и не обладает большой точностью прибор В. Р. Алеева (рис. 71), основанный на принципе условных угловых масштабов. При пользовании прибором Алеева рисовальный аппарат не требуется (Алеев, 1937). Угловой масштаб, т. е. масштаб с переменной единицей измерения, состоит из прозрачной пластинки (стеклянной или целлулоидной), на которую нанесен ряд радиусов, проведенных через равные углы. Перпендикулярно к среднему радиусу на протяжении всего масштаба нарезается ряд параллельных линий на расстоянии 0,5 мм одна от другой. В этом и состоит все устройство углового масштаба для обратного расчисления роста рыб по чешуе.
Угловые расстояния между смежными радиусами делают в 30 или 15 мин при длине радиуса в 5-6 мм.
Применять угловой масштаб можно двумя способами: 1) непосредственно накладывая масштаб на чешую, лежащую на предметном стекле и 2) вставляя его вместо окуляр-микрометра.
Величину масштаба непосредственно накладываемого на чешую целесообразно делать не больше, чем размер предметного стекла (77Х26,5 мм), в этом случае всего удобнее радиусы нарезать через ЗО’. Большинство рыб, для которых приходится обратно расчислять рост, бывают длиной не более 50-60 см. Поэтому, принимая расстояние между смежными радиусами по данному перпендикуляру за 1 см, достаточно нанести 50-60 радиусов.
Часто чешуя рыб, особенно толстая и крупная, не наклеивается, а прикрывается сверху стеклом, в этом случае более удовлетворяет своему назначению масштаб, вставленный в окуляр, но ввиду того, что для совмещения с размером изображения чешуи определенного числа радиусов необходимо масштаб передвигать, его нельзя делать круглым, как окуляр-микрометр. Делают две прорези в окуляре и вставляют масштаб на диафрагму. Для работы более удобны слабые окуляры (I-II).
Для быстроты отсчета каждый пятый радиус необходимо сделать толще (более резкой линией), чтобы он выделялся, а если технически возможно, то обозначать цифрами десятки радиусов через 1 см по длине масштаба.
Диаметр отверстия диафрагмы окуляра микроскопа равняется 10-11 мм, этим определяется ширина окулярного масштаба.
Окуляр вставляется в тубус микроскопа до верхней линзы, а так как окуляр с масштабом нельзя опустить ниже диафрагмы, то для удержания окуляра в таком положении на окуляр с прорезями нужно надеть медное пружинящее кольцо. Во избежание движения стекла с масштабом во время работы в трубке окуляра необходимо приделать пружинку, которая прижимала бы стекло к диафрагме.
В обоих вариантах масштаба для экономии места первый радиус удобней расположить вдоль края пластинки.
Применение крестообразного Столика для плавного передвижения чешуи на предметном стекле облегчает установку препарата. Изготовление столика нашими заводами уже освоено, но при работах с чешуей столик не обязателен.
Более подробное описание прибора В. Р. Алеева можно найти у Е. К. Суворова (1948). Н. И. Чугунова (1959) приводит описание прибора Брюзгина (рис. 72). Окуляр-масштаб Брюзгина также построен по принципу условного масштаба. Прибор представляет металлическую рамку длиной 12 см и шириной 17 мм; в рамку вводится прозрачная пластинка со шкалой на 500 делений, приготовленной делительной машиной или фотографическим путем (в последнем случае получаются расплывчатые линии и цифры). Сверху пластинки протягивается нить- волок или шелковая нитка; один ее конец закреплен неподвижно к раме, другой-к ползуну; передвигая последний, можно изменять угол а между нитью и линией, проходящей по нижнему краю пластинки.
Окуляр-масштаб вставляют в окуляр через :прорезь на уровне диафрагмы. Поперек диафрагмы натягивается тонкая нить – увязка-перпендикулярно к линии масштаба. При работе пользуются слабым объективом I. Линию указки совмещают по шкале с делением, соответствующим длине рыбы (в мм). Затем чешую передвигают так, чтобы ее центр совпал с линией пересечения указки и линии, проходящей по нижнему краю пластинки, после чего с помощью ползунка нить перемещают до совпадения с точкой пересечения края чешуи и указки, а ползунок закрепляют неподвижно. Установив масштаб, как указано, его передвигают до пересечения нити с краем годичных колец на линии указки; деление на шкале дает сразу искомый результат (l 1, l2, l3 и т. д.).
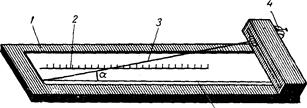
Рис 72 Окуляр-масштаб Брюзгина (по Чугуновой, 1959):
1-металлическая рамка; 2- прозрачная пластинка; 5-нить; 4- ползунок; 5-линия на краю пластинки с делениями; ?- угол между нитью (1) и линией (5).
Недостаток прибора – крупная чешуя карпа, леща и прочих не помещается под прибором.
В последнее время при определении возраста и темпа роста рыб стали применять различные проекционные аппараты что в значительной степени облегчает технику самой работы. К сожалению многие удачно изготовленные модели микроувеличителей построены кустарным способом и не имеют пока серийного производства. Из выпускаемых промышленностью проекционных аппаратов для этой цели используется обыкновенный фильмоскоп, приспособленный к штативу для вертикального проектирования. Для уменьшения фокусного расстояния Г. Г. Галкин (ГосНИОРХ) предлагает заменять линзу объектива фильмоскопа – окуляром 20 X, взятым от обычной школьной штативной лупы. Вторым прибором является микрофот типа 5 ПО-1, изготовляемый Московским киноэлектромеханическим заводом. Увеличенное изображение чешуи на экране расчисляется так же, как на доске Э. Леа.
Ю. Н. Редкозубов (1964) описывает микропроектор, сконструированный им путем использования микроскопа МБИ-1 и осветителя ОИ-19. Такой микропроектор очень удобен для определения возраста рыб по чешуе и срезам лучей плавников, а также для фотографирования препаратов. Изготовление его несложно. Используемые приборы не подвергаются переделке и в случае необходимости легко могут быть приведены в первоначальное состояние.
Анализируя различные методы реконструкции роста рыб по чешуе, Ф. И. Вовк (1955) пришел к заключению, что зависимость между длиной рыбы и размером чешуи не является прямолинейной, не точно укладывается она и в рамки параболической (Шериф, 1922) и логарифмической (Монастырский, 1930) связи. Отношения между длиной рыбы и размером чешуи изменяются с ростом рыбы всегда криволинейно. Связь между ростом рыбы и чешуи специфична для каждого вида, является его экстерьерным признаком и зависит прежде всего от строения чешуи, габитуса рыбы и изменения последнего в процессе индивидуального развития. На этом основании Ф. И. Вовк предложил новый метод расчисления роста рыб по чешуе с помощью предварительно составленных номограмм.
Подробное описание этого метода и техники построения номограмм имеется в работе Н. И. Чугуновой (1959).
Этот метод пока еще новый и нуждается в подтверждении на большом эмпирическом материале.
В последнее время у ихтиологов более широкое распространение получил метод изучения линейного и весового роста рыб по непосредственно определяемым величинам. Этот более простой метод дает практически удовлетворительные результаты при условии соблюдения сроков сбора материала, приуроченных ко времени завершения годовых циклов роста рыб.
При изучении роста рыб по непосредственно определяемым данным важное значение, как это отмечалось выше, имеет время сбора материала. По этому поводу П. В. Тюрин (1963) пишет следующее: «Рыба растет в основном летом; поздней осенью, зимой и весной рост у большинства рыб так же, как и частей, их тела, по которым определяется возраст, приостанавливается. Следовательно, материалы для определения возраста рыб выгоднее всего собирать в такой сезон, когда для характеристики линейного и весового роста по отдельным возрастным группам достаточно определяемых данных и отпадает надобность в обратных расчислениях. В то же время надо стремиться к тому, чтобы собранные материалы возможно достовернее отражали возрастную структуру промысловой части популяции изучаемого вида, что особенно важно для оценки запасов. В связи с этим большинство исследователей лучшим временем для сбора возрастных материалов считают период преднерестовых скоплений (для весенненерестующих рыб – весна и раннее лето, для осенненерестующих рыб – осень и ранняя зима).
Однако во внутренних водоемах промысел в преднерестовый и .нерестовый периоды нередко развит крайне слабо или запрещен правилами рыболовства. В таких случаях сборы возрастного материала следует приурочить к периоду основного промысла.
Так как рост рыб практически заканчивается ранней осенью, то материалы, собранные поздней осенью (при температуре воды ниже 6-4°С), можно сравнивать с материалами, полученными весной».
При сборе возрастных материалов, если уловы большие, берут среднюю пробу. Если рыбы добыто мало, измеряют весь улов. Эти материалы необходимы для выяснения возрастной структуры облавливаемой популяции. Для более полной характеристики возраста и роста рыб при сборе материала применяют выборочный метод, стараясь, по возможности, обеспечить наличие в сборах рыб всех возрастных групп, от молоди до старых особей включительно.
При обработке возрастного материала полезно пользоваться карточками, на которые записываются все полученные данные, относящиеся к изучаемой рыбе. Это позволяет быстро распределить материал по возрасту, длине, весу, полу, стадиям зрелости и т. п. По каждой возрастной группе вычисляют среднюю длину и средний вес, пользуясь, где это позволяет количество материала, методом вариационной статистики. Окончательные результаты обработки материалов по возрасту и росту рыб оформляются в виде таблиц. В качестве примера приведем в сокращенном виде таблицу из работы П. В. Тюрина (1957), в ней показаны средние размеры (длина до С) и вес леща из трех озер, полученные на основании наблюдавшихся данных (табл. 7).
Остановимся на способах вычисления средних размеров (по длине тела) отдельных возрастных групп и вычисления средних годовых приростов рыб. Однообразия в этих вычислениях нет, когда сравнивают быстроту роста рыб одного и того же вида по данным различно высчитанных средних, получаются выводы, не соответствующие действительности. Одни авторы среднюю возрастную длину определяют как среднее арифметическое из длин всех отдельных особей данного возраста; другие-общую среднюю возрастную длину находят, разбив исследуемых рыб первоначально по годам их выхода из икры.
Таблица 7
|
Озера |
Дата сбора материала |
Число экз. |
Возраст рыб |
||||||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
||||||
|
Длина, см |
|||||||||||||||
|
Велье |
V, начало VI |
154 |
7,4 |
13,6 |
20,6 |
26,6 |
30,8 |
35,2 |
37,0 |
39,1 |
42,1 |
– |
|||
|
Городенское Ильмень |
Средина VI V, начало VI |
181 548 |
6,55,4 |
13,1 9,6 |
19,914,4 |
25,1 18,6 |
28,1 22,4 |
31,2 27,7 |
34,5 32,6 |
37,1 35,0 |
39,5 37,9 |
– 38,9 |
|||
|
Вес, г |
|||||||||||||||
|
Велье |
7 |
49 |
189 |
395 |
637 |
1020 |
1151 |
1405 |
1664 |
– |
|||||
|
Городенсксе |
– |
44 |
193 |
303 |
458 |
641 |
823 |
1030 |
1240 |
– |
|||||
|
Ильмень |
2,5 |
15,2 |
57 |
140 |
241 |
438 |
735 |
936 |
1233 |
1338 |
|||||
Еще более разнообразны методы вычисления годовых приростов. Некоторые исследователи считают наиболее правильным способ нахождения средних возрастных длин, если эти возрастные средние определяются из средних длин отдельных возрастных групп. При небольшом количестве материала, взятого для определения темпа роста по методу обратного расчисления, наглядны таблицы, которыми пользуются многие исследователи (табл.8).
При изучении роста рыб важно знать, помимо линейных, весовые размеры рыб. Многими авторами, особенно работами П. В. Тюрина (1927), доказано, что вес рыбы находится, в прямой зависимости от длины тела.
Наглядный вид имеют и таблицы роста (линейного и весового) рыб, составляемые по примеру Ф. Д. Великохатько (1941). Одну из таких таблиц по темпу роста днепровского леща приводим ниже (табл. 9).
Есть различные способы вычисления годовых приростов. Прирост по длине тела чаще определяется путем вычитания длины тела, свойственной предыдущему году, из длины тела, свойственной последующему году.
Весовые, приросты некоторые авторы, например М. П. Борзенко (1935), выражают процентным отношением веса последующего года к весу предыдущего, причем средний вес годовалой рыбы в данном случае принимается за 100%.
Слишком мало сведений о росте молоди рыб в первые месяцы жизни, хотя они имеют огромное значение; темп ростa молоди (сеголетков) представляет хороший материал для изучения влияния различных гидрологических (температура, вый и солевой состав воды и т. д.) и гидробиологических (пищевых) факторов на рост молодого организма, особо чувствительного к этим факторам; темп роста сеголетков оказывает сильное влияние на дальнейший рост старших возрастных групп (годовиков, двухлетков и т.д.).
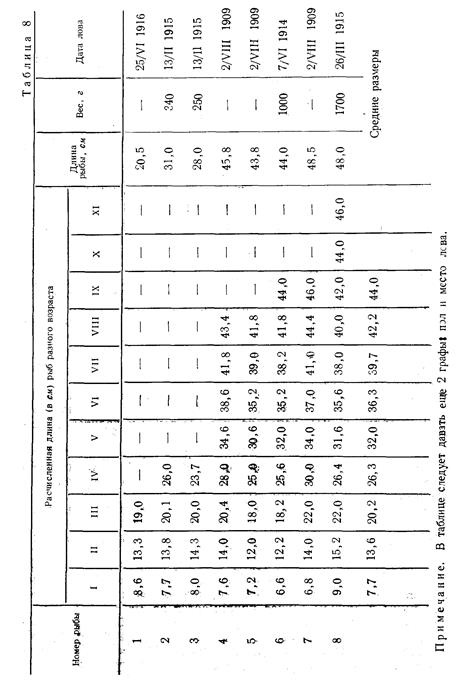
Таблица 9
|
Возраст |
Длина, мм |
Прирост, мм |
Вес, г |
Прирост, г |
Коэффициент упитанности |
Число рыб |
||||||
|
Самки |
||||||||||||
|
I |
96 |
96 |
18 |
18 |
– |
22 |
||||||
|
II |
153 |
57 |
78 |
60 |
– |
26 |
||||||
|
III |
245 |
92 |
326 |
248 |
2,22 |
91 |
||||||
|
IV |
301 |
56 |
573 |
247 |
2,10 |
259 |
||||||
|
V |
351 |
50 |
951 |
378 |
2,19 |
212 |
||||||
|
VI |
391 |
40 |
1315 |
364 |
2,20 |
296 |
||||||
|
VII |
421 |
30 |
1639 |
324 |
2,19 |
106 |
||||||
|
VIII |
449 |
28 |
1924 |
285 |
2,12 |
34 |
||||||
|
IX |
480 |
31 |
2335 |
411 |
2,11 |
14 |
||||||
|
X |
501 |
21 |
2512 |
177 |
2,23 |
11 |
||||||
|
XI |
520 |
19 |
– |
– |
– |
2 |
||||||
|
XII |
5ЭО |
10 |
– |
– |
– |
1 |
||||||
|
Самцы |
||||||||||||
|
I |
96 |
96 |
18 |
18 |
– |
22 |
||||||
|
II |
153 |
157 |
78 |
60 |
– |
26 |
||||||
|
III |
232 |
79 |
282 |
204 |
2,12 |
42 |
||||||
|
IV |
288 |
56 |
528 |
246 |
2,21 |
94 |
||||||
|
V |
343 |
55 |
816 |
288 |
2,02 |
208 |
||||||
|
VI |
381 |
38 |
1138 |
322 |
2,06 |
263 |
||||||
|
VII |
410 |
29 |
1416 |
278 |
2,05 |
41 |
||||||
|
VIII |
436 |
26 |
1647 |
23Г |
2,00 |
11 |
||||||
|
IX |
450 |
14 |
– |
– |
– |
1 |
||||||
|
Х |
460 |
10 |
– |
– |
— |
1 |
||||||
Обычно размеры годовиков сильно разнится даже в одной водоеме (если сборы рыб данного вида берут в различные годы)1. Подобная разница отмечается и у старших возрастных групп, но там она, как правило, выражена менее резко. Длину тела мальков (брать всю длину) следует измерять в мм, а вес определять в г.
Определение возрастного состава рыб в уловах
При установлении возрастного состава уловов обычно пользуются средними пробами, co6ирая их в больших количествах (на протяжении всей путины через 2-3 дня берется проба не менее 100 экз. рыб, за путину 10-12 проб, т. е. не менее 1000 рыб на каждом наблюдательном пункте). После определения возраста устанавливают процентные отношения полов в каждой возрастной группе. При большом материале рекомендуется определение возраста делать выборочно, но с таким расчетом, чтобы выборочные пробы имели все возрастные группы, например, берется четвертая часть общей большой пробы без выбора. А. В. Морозов предлагает в таких случаях пользоваться весовыми соотношениями, «чтобы иметь представление о том, в каких соотношениях представлены разные возрасты в уловах данного года, и на какие количества по весу мы можем рассчитывать на следующий операционный год» (Морозов, 1934).
Для перехода от счета к весу и обратно нужно иметь данные по соотношению между длиной и весом. Материал разбивают по длинам тела (например, через 1 см), определяют средний вес каждой такой группы. Например, получим средний вес для рыб в 15 см-65,4 г, для рыб в 20 см-161,8 г и т. д. Далее нетрудно подсчитать общий вес всех 15, 20-сантиметровых и других рыб. Полученные данные наносят на график с логарифмическими шкалами, такой график позволит рассчитать теоретические веса для тех размеров, которых в пробе нет. Таблицы соотношений длин и веса нужно составлять отдельно для самцов и самок.
Средние размеры некоторых рыб по возрастным группам
В табл. 10 указываются размеры ряда рыб, определенные по методу обратного расчисления.
Таблица 10
|
Возраст |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
|
Прирост |
13,7 |
11,2 |
12,0 |
11,0 |
11,0 |
10,6 |
11,3 |
10,3 |
9,1 |
9,1 |
8,5 |
5,7 |
Пользуясь этой таблицей, можно вычислить и годовые приросты рыбы за каждый год. Например, сом за первый год вырос на 13,7 см, за второй – на 24,9- 13,7=11,2 см. Поэтому легко составить и табличку годовых приростов аральского сома (табл. П).
Из этих данных видно, что наиболее быстро сом растет до трех лет, а затем годовые приросты постепенно уменьшаются. По таблицам прироста можно установить возраст, когда рыбу наиболее рационально вылавливать. Однако нужно знать что нарастание веса рыбы происходит и после того, как рыба закончила свой рост в длину. Расхождение приростов рыбы в длину и приростов веса хорошо показано на рис. 73.
Рис. 73. Сравнение приростов длины и веса воблы (по Правдину, 1939):
1- прирост длины; 2 – прирост веса.
Методы определения скрытых (потенциальных) возможностей роста рыб
Несмотря на исключительное разнообразие характера роста у различных видов и разновидностей рыб (подвидов, рас, племени, экотипов), есть основания для выделения нескольких основных типов линейного роста у рыб. Каждому из них свойственны свои особенности, которые заключаются в следующем.
I. Интенсивный рост в неполовозрелом возрасте с постепенным затуханием в половозрелом периоде жизни. ( Такой тип роста следовало бы считать наиболее важным, так как по И. И. Шмальгаузену (1935): «Скорость роста падает пропорционально времени, протекшему от его начала, обратно пропорционально возрасту».)
Этот тип роста чаще всего свойственен пелагическим рыбам с укороченным периодом жизни, например многим видам сельдевых, снетку, корюшке, ряпушке, рипусу, шемае, уклее, верховке, быстрянке (рис. 74).
II. Замедленный рост в неполовозрелом возрасте с заметным увеличением в начальный период половозрелой жизни и с угасанием к концу жизни.
Этот тип роста довольно часто встречается у бентофагов и хищников, например у сазана, леща, язя, осетра, тайменя, ленка, нельмы, щуки, сома, налима, судака и др.

Рис. 74, Чешуя рыбы с нормальным ростом, схема (по Дрягину, 1948).
Рис. 75. Чешуя рыбы с замедленным ростом в период речной жизни и интенсивным ростом в морской период, схема (по . Дрягину 1948).
Как вариант II типа роста – замедленный рост молоди в речной период жизни у проходных рыб и интенсивный в морской период жизни до достижения половозрелости с полным прекращением роста после первого же нереста вследствие гибели производителей (как у дальневосточных лососей и миног) или же с продолжением его в период половозрелой жизни (как у семги и кумжи) (рис. 75).
III. Почти равномерный тип роста как в неполовозрелом, так и в половозрелом возрасте без заметного изменения темпа роста после наступления половозрелости. Такой тип роста характерен для карликовых и камышовых форм, для многих видов прибрежных рыб, например у прибрежного окуня, плотвы, густеры, камышового леща, красноперки, карася, карликовой пеляди (рис. 76).
 Рис. 76. Чешуя рыбы с почти равномерным ростом, схема (по Дрягину, 1948).
Рис. 76. Чешуя рыбы с почти равномерным ростом, схема (по Дрягину, 1948).
IV.Неустойчивый, нерегулярный иногда скачкообразный тип роста, когда в неполовозрелом возрасте или в половозрелом, реже в том и другом, темп роста то ускоряется, то замедляется (рис. 77).
Для наглядного описания разнотипного хода линейного роста у рыб может быть применена так называемая характеристика роста по его удельной скорости, рекомендованная В. В. Васнецовым (1953а).
![]() Те же типы, как и в линейном росте, проявляются и в весовом приросте рыб, с той лишь особенностью, что с увеличением размеров рыб и возрастанием массы тела прирост веса (относительно к единице линейного прироста) планомерно увеличивается. По уклонению линейного и весового роста рыб часто можно судить о скрытых (потенциальных) возможностях роста того или иного вида рыб и отдельных популяций. При этом, следуя данным Е. К. Суворова (1940), под потенциальным ростом надо понимать «достижение при унаследованной конституции и при наличии благоприятных условий наибольших возможных результатов роста». Следует иметь в виду, что в природных условиях у многих рыб потенциальные возможности роста не реализуются в полной мере в результате неблагоприятных условий и неправильного ведения рыболовства. Имеется много доказательств, свидетельствующих о возможностях направленного воздействия на величину прироста и ускорение роста рыб путем регулирования плотности популяции, обогащения кормовой базы, улучшения газового режима и путем вселения рыб в более благоприятные условия. Приведем несколько примеров изменчивости в росте рыб.
Те же типы, как и в линейном росте, проявляются и в весовом приросте рыб, с той лишь особенностью, что с увеличением размеров рыб и возрастанием массы тела прирост веса (относительно к единице линейного прироста) планомерно увеличивается. По уклонению линейного и весового роста рыб часто можно судить о скрытых (потенциальных) возможностях роста того или иного вида рыб и отдельных популяций. При этом, следуя данным Е. К. Суворова (1940), под потенциальным ростом надо понимать «достижение при унаследованной конституции и при наличии благоприятных условий наибольших возможных результатов роста». Следует иметь в виду, что в природных условиях у многих рыб потенциальные возможности роста не реализуются в полной мере в результате неблагоприятных условий и неправильного ведения рыболовства. Имеется много доказательств, свидетельствующих о возможностях направленного воздействия на величину прироста и ускорение роста рыб путем регулирования плотности популяции, обогащения кормовой базы, улучшения газового режима и путем вселения рыб в более благоприятные условия. Приведем несколько примеров изменчивости в росте рыб.
Рис. 77. Чешуя пеляди со скачкообразным типом роста.
Длина рыбы по Смитту 395 мм, вес 1080 г, возраст 6 лет.
Река Колыма, 16/VI-1929 (по Дрягину, 1948).
По А. Н. Елеонскому (1946), у прудового карпа наблюдались следующие колебания веса в г:
Однолетние …..…….. 15-500
Двухлетние .….…….. 150-1000
Трехлетние …..……… 350-1500
Четырехлетние …….. 1000-3000
В прудах при различной плотности посадки отмечается различный вес сеголетков карпа. В прудах Молдавии при плотности посадки 2500 экз/га средний вес сеголетков составил 450 г, при 2000 экз/га- 625 г и при 1500 экз/га –1100 г.
По Е. К. Суворову (1940), при посадке в Балтийское море двухлетков ручьевой форели весом 70-80 г их вес увеличился за одно лето до 450-900 г и один экземпляр был весом 1140 г. Вес радужной форели за лето увеличился с 80 до 440-1100г.
По 3. П. Терещенко (1950), акклиматизированный в Каспии сингиль достигает 55 см длины и 2,5 кг весом, превосходя черноморского сингиля по весу в 3 раза.
По М. А. Фортунатову (1927), акклиматизированные сиги в озере Севане за 39 месяцев вырастали: лудога до 1200 г и чудской сиг до 1820 г, тогда как для такого прироста веса в материнских водоемах требуется около 8 лет.
По Б. П. Лужину (1956), акклиматизированная в оз. Иссык-Куль форель гегаркуни в возрасте 6+ имеет средний вес 3168 г (самцы) и 3882 г (самки), а в озере Севан соответственно 642 и 685 г.
Подобных примеров в литературе имеется очень много. Все данные по изменчивости роста у рыб указывают, что в природных условиях редко реализуются все возможности роста. Чаще всего можно заметить определенные признаки угнетения, сказывающиеся в задержке роста, особенно у популяций рыб с так называемым равномерным типом роста. Возникает вопрос, достигают ли рыбы потенциальных размеров и если достигают, то при каких условиях.
При изучении наибольших размеров рыб в природных условиях следовало бы выделять три группы, имеющие размер:
а) высокий, соответствующий потенциальному (обычен у осетровых рыб и часто у пелагических, а также хищных рыб);
б) депрессивный (у всех камышовых популяций рыб) и
в) резко пониженный (например, у сиговых рыб в бассейнах Колымы и Лены).
Основанием для биологической оценки наибольших размеров у рыб является закономерность, установленная впервые Фультоном (Fulton, 1906): «Рыбы приблизительно удваивают свою величину и их вес возрастает приблизительно в восемь раз после того, как они достигнут половой зрелости, или что рыбы приобретают половую зрелость, когда они достигнут около половины своей максимальной длины и около восьмой части своего максимального веса». Ряд примеров, подтверждающих положение Фультона, что наибольшие размеры самцов и самок в нерестовом стаде у рыб обычно в два раза превышают наименьшие, приведены в работе П. А. Дрягина (1934).
Эта закономерность наблюдается в разных группах рыб и рыбообразных: у миног, дальневосточных лососей, сельдей, карповых и других, и она проявляется как у рыб моноцикличных, так и полицикличных. При таком положении особенно важно учитывать уклонения от данной закономерности и выяснять причины их возникновения, а также искать возможности для воздействия на рыб для достижения ими потенциально возможных наибольших размеров.
Уклонения фактических наибольших размеров рыб от теоретических показаны на примере сиговых рыб Лены (табл. 12, по Дрягину, 1947).
Таблица 12
|
Вид |
Пол |
Наименьшая длина, см |
Средняя** |
Наибольшая •• |
|
Муксун |
самцы самки |
48 |
56/около 70 |
64/96 |
|
50 |
50/около 58 |
74/100 |
||
|
Омуль |
самцы самки |
41 |
48/около 60 |
56/82 |
|
74 |
52/около 66 |
60/88 |
||
|
Ряпушка сибирская |
самцы и самки |
24 |
30/около 36 |
35/48 |
|
Чир* |
самки |
41 |
50/около 60 |
58/82 |
* Река Колыма.
** Цифры слева – величины фактические, справа – теоретические.
На основании ряда опытных работ, связанных больше всего с переселениями рыб, установлено, что некоторые популяции с резко пониженными потенциальными размерами в новых водоемах достигают теоретически возможных размеров. Например, половозрелый рипус в Ладожском озере фактически бывает лишь длиною от 18-20 до 32 см, а в озере Увильды он вырастает до 39 см (Померанцев, 1949). Подобное же увеличение размеров отмечено у форели гегаркуни в Иссык-Куле и кефали сингиля в Каспии.
Таким образом, в размерах и росте рыб есть следующие закономерности, значение которых еще полностью не вскрыто и которые подлежат дальнейшему исследованию.
1. Наибольшая длина и продолжительность жизни рыб зависят от размеров и возраста наступления половозрелости.
2. Рыбы способны расти не только в неполовозрелом возрасте, но и в период половозрелой жизни. Разнотипность роста и размеров проявляются не только у отдельных видов, но и у отдельных рас и биологических групп.
3. Ограниченность и депрессивность в росте и наибольших размерах (потенциально свойственных данному виду) вследствие неблагоприятных условий питания.
4. Изменчивость в росте и размерах рыб в зависимости от условий обитания, воздействия промысла и численности стада.
5. Резко выраженная сезонность в росте со сменой периодов интенсивного роста периодами задержки или даже прекращения его на более или менее длительное время (иногда свыше полгода), часто в условиях полного голодания, при котором бывают большие потери в весе.
6. Случаи крайнего истощения при длительном голодании в условиях продолжающейся активной деятельности, связанной с нерестовой миграцией на большие расстояния и с нерестом, как это наблюдается у дальневосточных лососей, миног и угря.
В заключение следует отметить, во-первых, что недостаточно используется для ускорения оборота рыбного хозяйства исключительная пластичность в росте и размерах рыб под воздействием улучшения условий их обитания и регулирования численности стад и, во-вторых, при изучении роста и развития рыб надо направить основное внимание на выявление специфичности роста и развития в отдельные фазы жизненного цикла рыб, их взаимозависимости и взаимодействия.
Определение упитанности и жирности рыб
Для определения степени упитанности рыб широко пользуются коэффициентом Фультона, вычисляемым по формуле
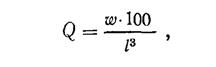
где Q – коэффициент упитанности;
w – вес рыбы, г;
l – длина рыбы от начала рыла до конца чешуйного покрова, см.
При определении коэффициента упитанности берется общий вес рыбы (вместе со всеми внутренностями). Такой способ далеко не всегда отображает истинные показатели упитанности. Различная степень развития половых продуктов и наполнения кишечника мешают нахождению правильного коэффициента упитанности. Например, при вычислении коэффициента упитанности горбуши с момента подхода ее к устью Амура и до нереста коэффициент упитанности обнаружил тенденцию к увеличению по мере приближения горбуши к нересту. Между тем и без вычислений было ясно, что упитанность рыбы падала. Увеличение коэффициента здесь произошло вследствие увеличения веса половых продуктов горбуши. Удлинение тела рыбы за счет преднерестового и нерестового увеличения длины рыла в этом опыте не повлияло на коэффициент упитанности, потому что я брал длину тела от жаберной щели до конца чешуйного покрова, т. е. длину туловища.
При вычислении коэффициента упитанности следует пользоваться длиной туловища, так как именно эта часть наиболее характеризует упитанность рыбы.
Более показательные результаты дает коэффициент упитанности по Кларк, вычисляемый по весу рыбы без внутренностей. Г. В. Никольский рекомендует пользоваться одновременно обоими способами.
Все же ни тот, ни другой коэффициент упитанности не дают каких-либо определенных количественных показателей упитанности, а только сравнительную оценку упитанности.
В рыбоводной практике, особенно в карповодстве, сравнительная оценка упитанности рыб производится путем определения отношения высоты тела к ее длине, чем больше это отношение, тем карп считается упитаннее, т. е. упитанность определяется высотой тела. Такой коэффициент упитанности менее надежен, чем коэффициенты Фультона и Кларк, так как и при очень высоком теле рыба может иметь низкую упитанность.
Периодическое определение степени упитанности имеет существенное значение при прогнозах срока икрометания рыб, так как чем выше коэффициент упитанности, тем скорее следует ожидать наступления нереста. В. В. Покровский подметил, например, что у ряпушки в Онежском озере массовый нерест может быть и при температуре 5-6 и при 2-3 °С, и что срок нереста во многом зависит от степени упитанности нерестующих рыб. При хороших условиях летнего питания ряпушки коэффициент упитанности выше, половые продукты созревают быстрей, и это ускоряет наступление нереста.
В. А. Амосовым (1961) разработана методика определения нового показателя экстерьера и упитанности рыб, не зависящего от степени развития половых желез, наполнения пищеварительного тракта и других факторов и названного автором «индексом удельной вальковатости».
Метод В. А. Амосова основан на изучении формы тела рыб в поперечном плане, в частности на определении поперечных контуров спины рыб в месте наибольшей высоты их тела. Автор’ указывает, что спинная часть рыбы является основным филейным участком, она более постоянна по форме, чем брюшная часть, и хорошо характеризует основное хозяйственное качество рыбы – мясистость.
Основные измерения, рекомендуемые автором, следующие: длина рыбы до конца чешуйного покрова, длина головы, наибольшая высота тела, наибольшая высота спины, измерения в поперечном сечении по трем характерным точкам полудуги спины. Последние измерения производятся при помощи специально сконструированного В. А. Амосовым прибора – полярно-координатного циркуля. Автор предлагает также эмпирическую формулу для определения количества (веса) мяса рыбы.
Жирность рыб точно определить можно только путем химического анализа, и ихтиологи для характеристики этого показателя получают обычно готовые, данные от соответствующих специалистов.
При работах на местах пользуются упрощенными методами •определения степени жирности. М. Л. Прозоровской разработана следующая пятибалльная шкала для определения жирности воблы, которая может быть использована и при работах с .другими рыбами (по Никольскому, 1963).
Балл 0. Жира на кишечнике нет. Иногда кишечник покрыт тонкой белой соединительной пленкой. Между петлями кишечника видны нитевидные образования этой пленки.
Балл 1. Тонкая шнуровидная полоска жира расположена между вторым и третьим отделами кишечника. Иногда по верхнему краю второго отдела проходит очень узкая прерывающаяся полоска жира.
Балл 2. Неширокая полоска довольно плотного жира меж-.ду вторым и третьим отделами кишечника. По верхнему краю второго отдела идет узкая непрерывная полоска жира. По нижнему краю третьего отдела кое-где виден жир отдельными небольшими участками.
Балл 3. Широкая полоска жира в середине между вторым и третьим отделами кишечника. В петле между вторым и третьим отделами эта полоса расширяется. По верхнему краю второго отдела и нижнему краю третьего идут широкие жировые полосы. У первого изгиба кишечника, если считать от головного конца, имеется жировой вырост в виде треугольника. Анальный конец кишечника в подавляющем большинстве случаев залит тонким слоем жира.
Балл 4. Кишечник почти целиком покрыт жиром за исключением маленьких просветов, где видна кишка. Эти просветы обычно бывают на второй петле и на третьем отделе кишечника; иногда можно встретить такие просветы и на втором отделе. Жировые выросты на обеих петлях мощные.
Балл 5. Весь кишечник залит толстым слоем жира. Нет никаких просветов. Мощные жировые выросты на обеих петлях.
Глава IX
МИГРАЦИИ
Миграции – величественное по красоте и глубокое по биологическому смыслу явление в жизни рыб. Вместе с тем они имеют первостепенное значение для промысла: массовый лов многих рыб производится на путях передвижений их стад. Поэтому изучению миграций рыб уделяется большое внимание и этой проблеме посвящено неисчислимое количество заметок, статей, книг. Одной из лучших книг является упоминавшаяся уже монография П. Ю. Шмидта-«Миграции рыб» (1947). Книга обогащена сведениями о миграциях рыб, почерпнутыми из советской и иностранной литературы. Приступающие к исследованию миграций рыб должны хорошо проштудировать книгу П. Ю. Шмидта прежде, чем начать свои исследования. Известна также сводка о миграциях рыб английского исследователя Мика (Meek, 1916). Подробно описываются миграции рыб в книгах Е. К. Суворова (1948), Г. В. Никольского (1963} и в других руководствах по ихтиологии.
Классификация миграций и их биологическое значение
Массовые передвижения рыб вызываются потребностями их организма в разные периоды жизненного цикла и сезоны. Связаны они главным образом с размножением, питанием и зимовкой.
Передвижения рыб к местам размножения носят название нерестовых миграций. Лососи из морей или озер идут размножаться в реки, иногда на огромные расстояния. Атлантический угорь, наоборот, из пресных вод направляется на нерестилища, расположенные в океане, за тысячи километров от мест нагула.
Рыбы, передвигающиеся в поисках пищи, совершают кормовые миграции. Например, исхудавшая после нереста у берегов Норвегии треска направляется к востоку, в мурманские воды, где находит хорошие пастбища.
Многие рыбы на зиму уходят в более глубокие участки водоема, такие миграции называются, зимовальными. Они свойственны некоторым рыбам Северного Каспия, массами залегающим на ямах. Хамса после нагула в Азовском море уходит на зиму в Черное море.
Передвижения рыб в толще воды называются вертикальными миграциями. Они чаще носят характер кормовых миграций, обусловливаются передвижениями кормовых объектов и хорошо выражены у планктоноядных рыб (ряпушка, салака и др.).
Икра и личинки многих рыб, особенно морских, а также нерестящихся в реках, переносятся на большие расстояния течениями. Это пассивные миграции, например, миграции икры и личинок атлантического угря из Саргассова моря к берегам Европы.
Передвижения рыб, которые кормятся в морях или озерах, а для размножения подходят к берегам или поднимаются в реки, называют анадромными миграциями, в отличие от катадромных миграций рыб, передвигающихся на нерест из рек в моря или от берегов в глубины открытого моря.
Употребляются иногда термины контранадромная миграция(передвижение против течения) и денантная миграция(сплывание по течению).
На основании характера миграций строятся биологические классификации рыб.
К. Ф. Кесслер (1877) предложил делить рыб на морских, солоноват сводных, разноводных, проходных, полу проходных и пресноводных. После К. Ф. Кесслера предлагались и иные биологические классификации: рыб, но в основе всех их лежит схема Кесслера.
Е. В. Бурмакин и П. В. Тюрин (1959) после критического обзора прежних классификаций рыб, основанных на их .миграциях, предложили новую, более совершенную классификацию.
При делении рыб на биологические группы они принимают во внимание только те факторы, которые определяют среду обитания рыб в течение важнейших периодов их жизни, а именно – в периоды нагула и размножения.
«Если среда, в которой происходит основной период нагула рыбы, сходна со средой, где в основном происходит и естественное воспроизводство, то мы имеем местную или туводную форму. Если же среда в том и. другом случае не сходна и рыба для естественного воспроизводства или для нагула должна совершать миграцию из одной среды в другую, резко отличную (например, из морской или осолоненной воды в пресную речную или в обратном направлении, а также из озера в реку), то-это будет форма проходная – морская, солоновато-водная или пресноводная» (Бурмакин и Тюрин, 1959).
Полиморфные виды, указывают далее эти авторы, могут иметь формы (популяции), относящиеся к разным биологическим группам: одни к непроходным, другие к проходным. Такие формы свойственны, например, гольцам (Salvelinus), ряпушкам, сигам и многим другим видам. Это свидетельствует о процветании таких видов, расширяющих свои ареалы. В ниже помещаемой классификации виды рыб, приспособившиеся к обитанию в разной среде (полиморфные виды), отмечены звездочками.
Биологическая классификация рыб по Е. В. Бурмакину и П. В. Тюрину
Непроходные рыбы
1. Морские рыбы, нагуливающиеся и размножающиеся преимущественно в водах с морской соленостью и не совершающие миграций в реки. Примеры: сельдевая акула, полярная акула, тихоокеанская сардина, атлантическая сельдь, тихоокеанская сельдь, мойва, треска, пикша, сайда, морской окунь, тунец, палтус, камбала-ерш.
2. Рыбы солоноватоводные, нагуливающиеся и размножающиеся преимущественно в солоноватоводных участках морей или в солоноватых озерах (типа Арала и Каспия) и не совершающие миграций в реки. Примеры: тюлька, хамса, анчоусовидная килька, долгинская сельдь, салака, аральская вобла, морской судак, барабуля, бычок-кругляк, аральская шемая *, аральский лещ *, лобан, сингиль, ледовитоморская рогатка, полярная камбала.
3. Рыбы пресноводные, нагуливающиеся и размножающиеся преимущественно в пресных водах и не совершающие нерестовых миграций, связанных с резкой сменой среды. Могут быть разделены на три подгруппы:
а) озерные, предпочитающие стоячие воды. Примеры: палья, сиг лудога, снеток, озерный гольян, линь, карась, голец *, ряпушка *, пелядь*, балхашская маринка *.
б) речные, предпочитающие текучие воды. Примеры: лопатонос, таймень, ленок, тугун, валек, чукучан, елец, голавль, подуст, пескарь, калуга*, стерлядь*, сибирский осетр*, ручьевая форель, балхашская маринка *;
в) общепресноводные, живущие и нерестующие как в озерах, так и в реках. Примеры: щука, плотва, язь, красноперка, уклея, синец, окунь, ерш, пыжьян*, лещ*, чехонь*, сазан *, налим *, судак *.
4. Рыбы разноводвые, способные постоянно жить, нагуливаясь и размножаясь в пресных, солоноватых или морских водах. Пример: трехиглая колюшка.
Проходные рыбы
5. Морские проходные рыбы,нагуливающиеся в море, откуда для нереста они входят в реки и совершают по ним миграции. Примеры: кета, горбуша, чавыча, семга, восточная красноперка, кумжа, голец *.
6. Солоноватоводные проходные рыбы, нагуливающиеся в солоноватоводных участках морей или в солоноватых озерах (типа Арала и Каспия), откуда для нереста входят в реки и совершают по ним миграции. Примеры: каспийская минога, белуга, севрюга, черноспинка, черноморская сельдь, каспийский лосось, белорыбица, сибирская ряпушка, омуль, муксун, азиатская корюшка, тарань, каспийская вобла. кутум, черноморская шемая, каспийская шемая, рыбец, калуга *, сибирский осетр *, нельма *, ряпушка *, пыжьян *, корюшка *, лещ*, аральский лещ*, сырть *, чехонь*, сазан*, судак*.
7. Пресноводные проходные рыбы, нагуливающиеся в пресных озерах или пресных морских губах, откуда для нереста входят в реки и совершают по ним миграции. Примеры: озерный лосось, байкальский омуль, чир, волховский и сунской сиги, нельма*, пелядь*, пыжьян*, корюшка*, балхашская маринка *, сырть *, налим *.
8. Катадромные проходные рыбы, нагуливающиеся в пресных водах, откуда для нереста мигрируют по рекам в море. Пример: угорь.
Наиболее грандиозны по масштабам – по количеству особей и по протяженности передвижений-миграции морских проходных и солоноватоводных проходных рыб. К ним, в частности, относятся дальневосточные лососи (кета и горбуша), которые миллионами особей устремляются на нерест в Амур, Камчатку и другие реки Дальнего Востока. Многим морским рыбам, например сельдям и треске, также свойственны весьма длительные по времени и протяженности миграции, кормовые и нерестовые. Миграции пресноводных рыб не так значительны по количеству мигрирующих рыб и по протяженности самих миграций, что вполне понятно, если иметь в виду размеры морских и пресноводных водоемов.
Каково же биологическое значение миграций рыб? Миграции следует рассматривать как одно из приспособлений к увеличению численности вида, развивавшееся и закрепленное в процессе его эволюции.
При миграциях рыбы «активно меняют условия своего существования, используя наиболее выгодные стороны разных биотопов, избегая по возможности их невыгодных сторон» (Мантейфель, 1959). В пределах одного и того же вида значительно большая численность бывает у мигрирующих форм, чем у популяций с ограниченным ареалом.
Так, в Каспийском море имеется несколько форм бражниковской сельди (Alosa brashnikovi), но из них наибольшей численностью обладает «долгинка», совершающая миграции из Южного Каспия в Северный. Среди многих форм океанической сельди (Clupea harengus) самыми многочисленными являются атлантическая, норвежская и тихоокеанская сельди с очень большими по протяженности кормовыми миграциями (Никольский, 1963).
При таких миграциях рыбы полнее используют кормовые ресурсы водоемов, удлиняется и период их нагула. В этом отношении весьма показательные примеры приводятся Б. П. Мантейфелем (1959) для сельди и трески.
Атлантическая сельдь в Норвежском и Гренландском морях питается преимущественно рачками Calanus и Jhemisto в период их преднерестовых скоплений. Но эти скопления кратковременны и биомасса рачков, используемая в данном районе планктофагами, не восстанавливается. Однако в Норвежском и Гренландском морях размножение рачков и их преднерестовые скопления происходят не одновременно, а постепенно перемещаются в северном направлении. Соответственно этому основное стадо сельди также постепенно мигрирует на север, причем скорость его передвижения соответствует срокам преднерестовых скоплений рачков в разных районах. Мигрирующие сельди, говорит Б. П. Мантейфель, все время переходят с одних преднерестовых скоплений рачков Calanus и Themisto на другие, вновь образующиеся севернее предыдущих. В результате длительных миграций с юга на север сельдь значительно увеличивает период откорма.
Треска после зимовки и нереста .у Лофотенских островов мигрирует в южной части Баренцева моря в восточном направлении. С февраля по апрель она держится у мест зимовок мурманской сельди, питается также находящимися здесь мойвой и молодью тресковых рыб. В июне-июле треска появляется на склонах отмелей центральных районов Баренцева моря, где в это время в огромных количествах скопляется рачок «капшак» (Euphausiacea). Уничтожив капшака, треска передвигается в восточные районы моря, переходя на питание креветками и сайкой.
Таким образом, указывает Б. П. Мантейфель, треска использует ежегодно сроки и места скоплений наиболее массовых кормовых организмов. «Кормовая миграция, возникшая в процессе эволюции, как адаптация к использованию пищи при ее наибольшей доступности, отражает в себе особенности биологии кормовых организмов. Разнообразные внешние и внутренние сигналы миграций, вероятно, дают возможность мигрирующей популяции выдерживать соответствующие сроки, не опаздывая и не опережая сроков скоплений кормовых организмов» (Мантейфель, 1959).
Нерестовые миграции, согласно В. В. Васнецову (по Мантейфелю, 1959), следует рассматривать как защитную адаптацию от хищников, истребляющих икру и молодь рыб. Так рассматриваются миграции проходных рыб, откладывающих икру в реках с меньшей численностью рыб – хищников, чем в морях (или озерах). Нерестовые миграции «полупроходных» (солоноватоводных проходных) рыб, считает Б. П. Мантейфель, имеют и то значение, что их развивающаяся молодь оказывается в более благоприятных условиях в отношении обеспечения кормом в полоях (где нерестятся эти рыбы).
Методы изучения миграций
Исследования миграций рыб должны сопровождаться гидробиологическими, гидрологическими и метеорологическими исследованиями.
Наиболее подробно изучены миграции нерестовые, и методика их изучения освещена во многих работах достаточно полно. Например, Н. П. Танасийчук (1948) дал полную картину мест и сроков миграций волжской сельди в море, дельте и в нижних участках Волги на материале пятилетних наблюдений (1935-1939 гг.). Миграции он сопоставил с экологическими условиями (температурой воды, соленостью, ветрами и течениями). Дал характеристику нерестовых стад сельди и снабдил работу картами миграций сельди.
В основном наблюдения и сборы материалов по миграциям рыб производятся при работах на научно-исследовательских судах, а также на береговых пунктах, частично путем сбора анкетно-корреспондентских сведений. Все данные, собираемые на наблюдательных пунктах по улову ходовой рыбы, также освещают отдельные моменты миграций.
В пунктах, где промысел по тем или иным причинам мало интенсивен или совсем отсутствует, а также где между отдельными промысловыми сезонами существует длительный перерыв, целесообразно организовать собственный опытный лов с помощью сетных порядков, составляемых из сеток, имеющих разную по размерам ячею.
Инструкция Каспийской экспедиции рекомендует следующие правила, которые нужно выполнять при опытных ловах.
Количество выставляемых на каждом пункте опытных сетей, равно как и способы их установки, могут варьировать в зависимости от местных условий. Однако для большей четкости результатов опытного лова при расстановке сетей, опытных порядков необходимо придерживаться определенного плана:
1. Располагать опытные порядки на разном расстоянии от берега и на разных глубинах. При этом, как правило, следует принять, что средний порядок располагается в местах наиболее интенсивного для данного района промысла, бережной – ближе к берегу, а глубинный – за пределами обычной промысловой зоны.
2. Всюду, где это можно по гидрологическим условиям, порядки располагаются попарно в противоположных друг к другу направлениях, например один порядок перпендикулярно по отношению к берегу, другой на небольшом расстоянии от него – параллельно берегу, или один поперек течения, а рядом с ним выставляется порядок по течению.
3. Принятый порядок и места расстановки опытных сетей не менять без достаточных к тому оснований.
4. План и условия расстановки опытных порядков (расстояние от берега, расположение порядков в отношении последнего, направление порядков по длине и глубине места выбивки), а равно все последующие изменения и перемещения тщательным образом фиксировать в журналах, в дневниках и на рабочих картах.
При осмотре опытных порядков в целях установления направления движения рыбы наблюдают, с какой именно стороны преимущественно вошла в сеть рыба, а также какие размеры ячей являются для данного места наиболее уловистыми. Кроме того, при -каждой переборке порядков, а равно и смене их, производятся, по возможности, у каждого опытного порядка и обязательно у порядков бережного и глубинного, определения поверхностной температуры воды и направления течения. Все наблюдения заносятся в дневник или особый журнал.
При выборке выловленной опытными порядками рыбы надо сортировать ее для каждого порядка отдельно, не смешивая с рыбой других опытных порядков. Необходимо тщательно отмечать размер ячей сетей, составляющий сетной порядок, из которого выбрана рыба. Общий учет рыбы производится обязательно в средних показателях на сетку (каждого из выставленных размеров ячей) за ночь лова. Это является показателем данного улова.
Описанный порядок учета опытных уловов (для сбора сведений по миграции рыб) принят на промыслово-исследовательских пунктах Каспийского моря. Этот способ можно применять и в других местах, причем наблюдатель, несомненно, сумеет внести в приведенные указания свои дополнения и изменения, учитывая условия миграции тех рыб, над которыми ведутся наблюдения, а также морфометрические особенности водоемов.
При наблюдениях над миграцией миноги в нешироком рукаве Волги (Правдин, 1913), мною было установлено свойство миноги идти ночью и в туманные дни в верхних слоях воды, а днем и в светлые лунные ночи-в придонных.
Наблюдения над промысловым ловом рыбы помогают установлению сроков миграций. Появление рыб в неводах или в сетных уловах указывает на начало ее хода в данном месте.
Миграции рыб в реках могут быть установлены и непосредственными наблюдениями. Вошедший в реку каспийский осетр время от времени поднимается к поверхности воды и выпрыгивает из нее. У волжских рыбаков это называется «осетр, взмыл». Ход кеты на амурских притоках можно также наблюдать на мелких обычно приустьевых перекатах, рыба стремительно проходит через эти перекаты.
Подобные наблюдения могут быть и в открытых частях морей. На Охотском море против западного берега Камчатки я наблюдал миграцию горбуши, которая выпрыгивала из воды почти по вертикальной линии. Японские исследователи наблюдали ход кеты в море также по характеру ее прыжков, которые кета делает по дугообразной линии. Таким образом, можно различать рыб и их миграции и по характеру их прыжков.
Еще ярче выражены скопления мигрирующих пелагических рыб. Громаднейшие стаи сельдей мигрируют в верхнем горизонте воды в таких количествах, что их миграции можно видеть с палубы судна и еще лучше с самолета. Последний метод наблюдений над миграциями сельдей широко применяется на Дальнем Востоке. А. Г. Кагановский таким методом изучил поведение тихоокеанской сардины (иваси) и тем самым оказал существенную помощь промыслу. В книге Ю. Ю. Марта «Промысловая разведка» (1948) приводится много примеров, показывающих значение наблюдений с самолета над миграциями рыб. Так, установлено (с самолета), что иваси в зависимости от температуры воды изменяют форму своих косяков, их вертикальное расположение и мощность самих косяков. Температура, ветер, распределение и количество корма (планктона),- все это факторы, влияющие на миграции иваси.
А. Г. Кагановский (1951), изучая миграции скумбрии в Японском море, отметил две формы миграций-кормовую и нерестовую. По этому поводу А. Г. Кагановский пишет: «по мере потепления в северных, соседних с местами зимовки скумбрии в отепленных водах Цусимского пролива районах, скумбрия начинает мигрировать на север независимо от степени зрелости половых продуктов, при этом довольно интенсивно питается. Во время этой миграции половые продукты скумбрии созревают, и она нерестует в прибрежной зоне. Закончив нерест, скумбрия немедленно отходит с нерестилищ и продолжает миграцию в открытом море (кормовую). Осеннее похолодание вызывает обратную миграцию скумбрии на юг.
Слабо изучены суточные вертикальные перемещения, которые присущи многим рыбам (и пресноводным и морским) и также имеют решающее влияние на ход промысла.
Минога в светлые ночи идет у дна, а в темные поднимается в более высокие горизонты. Сиги Ладожского озера в светлые (белые) ночи также опускаются в более глубокие слой. Суточное вертикальное перемещение установлено К. Ф. Телегиным (1938) для салаки в Финском заливе. И. И. Николаев (1954) приводит данные свои и других исследователей, подтверждающие, что у салаки существуют суточные вертикальные миграции: в светлое время суток салака держится в более глубоких горизонтах моря, а в сумерки и ночью (даже и в лунные ночи И. П.) поднимается ближе к поверхности. Такое перемещение салаки автор ставит в прямую связь с вертикальными перемещениями главной пищи салаки (мизид и копепод).
Ихтиолог должен построить свои исследования так, чтобы вертикальные перемещения рыб были учтены и промыслом. В этом отношении полезно применять опытный так называемый ступенчатый лов рыбы путем установки орудий лова на разных глубинах.
Исследование вертикальных, сезонных и суточных миграций рыб необходимо вести совместно с изучением питания рыб и гидрометеорологических факторов.
Миграции хищных рыб обычно связаны с миграциями рыб, потребляемых этими хищниками. Ладожские лососи Весной идут следом за корюшкой. С. М. Кагановская (1937) пишет, что колючая акула (Squalus acanthias) ранней весной идет за косяками сельди, а после сопровождает косяки иваси.
При изучении рыб в период их миграций, особенно миграций нерестовых, нужно следить за группировкой рыб по возрасту, полу, расовым и другим отличиям. Известны так называемые первые, вторые и третьи подходы дальневосточной кеты. О мурманской сельди сообщается, что ее летние подходы к берегам Мурмана имеют три фазы: 1) в мае (иногда в июне) подходит мелкая летняя сельдь, 2) в июле (иногда в июне) подходит крупная сельдь и 3) в августе и сентябре идет типичная жирная сельдь (fatsild). В отдельные годы такая группировка несколько нарушается. Мелкая летняя сельдь бывает представлена преимущественно годовиками. Ихтиолог в подобных случаях должен вести анализ (главным образом биологический анализ) мигрирующих рыб по отдельным подвидам.
Подобные явления известны и для семги, которая в северные реки мигрирует стадами, разнящимися по возрасту и по биологии, но почти не отличающимся по морфологическим признакам (летняя семга, осенняя, тинда). У пресноводных рыб во время их миграций также наблюдаются возрастные группировки.
Наиболее слабо изучены миграции рыб после их нереста, «скатывание» рыб. Даже не установлены точно сроки ската, не говоря о его путях и о точных местах пребывания только что скатившейся рыбы. Трудность исследований такого рода миграций заключается, во-первых, в том, что скат многих рыб происходит подо льдом зимой и ранней весной, во-вторых, при скате не бывает массовых скоплений рыб и потому такие миграции обычно ускользают от внимания ихтиологов, хотя они имеют не меньшее биологическое значение, чем обычные массовые нерестовые миграции.
Нет обстоятельных данных и о том, как быстро восстанавливается производительная способность многих рыб после нереста, особенно рыб, нерестующих осенью.
Нужны наблюдения и над миграциями личинок и молоди: об отходе молоди с нерестилищ на пастбища, о миграциях ее из прибрежной зоны в более глубокие места, о скате молоди лососей и озерно-речных сигов из рек в моря и озера.
Некоторым рыбам свойственны особого рода миграции, которые невозможно считать кормовыми или нерестовыми. Нередко отмечаются случаи, когда вылавливают необычную для данной местности рыбу (сазан в реке Вятке, скумбрия в Белом море и т. п.). Стали учащаться ранее чрезвычайно редкие случаи захождения дальневосточных лососей (кеты) в сибирские реки. Каждый такой случай необходимо точно регистрировать, и старательно собирать сведения обо всех таких случаях (через местных рыбаков, краеведов, через местные органы печати). Подобные факты чрезвычайно важны не столько в биологическом отношении, как в отношении климатическом. И чаще такие случаи своим происхождением обязаны временным колебаниям температуры воды, которые распространяются на обширные водные пространства.
Изучение миграционных путей рыб дает также хорошие материалы и по истории ихтиофауны данного края. Ф. Д. Великохатько (1938), разобрав миграции многих рыб бассейна Черного моря и сопоставив пути миграций с геологическим прошлым этого бассейна, пришел к заключению, что филогенетически более старые рыбы связывают свои миграции с более старыми руслами поемных речек и их старицами. Это особенно ярко проявляется в миграциях осетровых рыб в дельте и нижнем течении Днепра. Филогенетически молодые виды рыб (например, тарань, вырезуб, рыбец) движутся по новейшим поемным магистралям Днепра.
Способы мечения рыб и образцы меток
Мечение рыб- один из самых надежных способов наблюдений для миграциями. Мечением можно пользоваться и для других исследований биологии рыб, например для наблюдений над ростом, над короткими и краткосрочными перемещениями, над выживаемостью рыб после нереста и т. п.
Процесс мечения рыб должен проводиться по хорошо разработанному плану, предусматривающему подготовку к мечению и само мечение. Имеется много указаний по мечению, есть и особая, составленная Г. А. Караваемым (1958), инструкция по мечению рыб.
До начала работ по мечению нужно возможно шире оповестить (специальными плакатами и через периодическую печать) о проводимом мечении. В плакате полезно давать рисунок метки и способ ее прикрепления к рыбе. Необходимо указывать точный адрес, куда направлять метки с пойманных рыб. Обычно назначаются вознаграждения за поимку меченой рыбы.
Работы по мечению регистрируют в особом журнале. Журнал, как указывается в инструкции Г. А. Караваева, должен содержать такие графы: 1) порядковый номер, 2) номер метки, 3) час и минуты, число, месяц и год мечения, 4) место выпуска, 5) длина рыбы, 6) вес, 7) особые отметки, 8) отметки о поимке вновь. Для каждого вида рыбы ведется особый журнал. От меченой рыбы берут несколько чешуек для определения возраста.
Мечение рыб приемами, которые могут вызывать длительный болезненный процесс, например мечение взрослых рыб отрезанием части жаберной крышки или одного из плавников не следует допускать.
Так как процесс мечения (измерение и взвешивание рыб, взятие чешуек, прокол тела, прикрепление метки) причиняет рыбе беспокойство и боль, рекомендуется поместить несколько меченых рыб на некоторое время в садок, чтобы проверить влияние мечения на поведение рыбы. Такую отсадку лучше делать до начала массового мечения.
Хорошая сводка о мечении рыб составлена Н. Е. Аслановой. В ее статье «Мечение промысловых рыб» (1961) говорится о развитии этого метода изучения миграций рыб в нашей стране, начиная с опытов Н. А. Бородина по мечению осетров в Волге и Каспийском море в 1871 г.
В советское время рыб метят на всех морях и многих пресноводных водоемах-реках, озерах и водохранилищах.
Благодаря этому методу изучены нерестовые, кормовые и зимовальные миграций ряда важнейших промысловых видов, например трески и пикши (Маслов, 1944; Константинов, ‘,1956, 1957), атлантической сельди (Марти, 1956; Вильсон, 1956), дальневосточной сельди (Дружинин, 1959), каспийской севрюги (Державин, 1922), воблы (Караваев, 1939), черноморской камбалы (Марти, 1940), ставриды (Сафьянова, 1958; Georguiev et Kolarov, 1959), кефали (Александрова, 1957) и других рыб.
Посредством мечения определялись также интенсивность промысла и состояние запасов рыб, например беломорской трески (Сонина, 1957), коэффициент уловистости орудий (Андреев, 1949; Иоганзен, 1952), промысловый возврат, совершенствовалась методика определения возраста и темпа роста рыб.
Советский Союз имеет взаимную информацию о мечении рыб с 18 странами мира. Международным Советом по исследованию морей печатаются, сообщения о способах мечения рыб, большие обзоры опубликованы в издании «Jornal du Conseil» в 1932 и 1953 гг. (Асланова, 1961).
В Инструкции по мечению рыб, составленной Г. А. Караваевым (1958), говорится, что метки должны удовлетворять следующим основным условиям: а) не наносить рыбе больших ран, которые могут привести к заболеваниям, а также будут мешать движению и другим жизненным функциям рыбы; б) быть легкими, прочными, стойкими к воздействию внешней среды, хорошо заметными, достаточно простыми и недорогими.
В изложении Г. А. Караваева ниже дается описание наиболее распространенных типов меток и способов мечения рыб (Караваев, 1958, стр. 21-37).
Индивидуальное мечение
При индивидуальном мечении каждая метка имеет свой порядковый номер.
Метки нумеруют или просто порядковым номером или сериями, т. е. буквой и порядковым номером, например, А-253, В-127. При мечении в открытых морях или в международных водах рекомендуется для обозначения серий употреблять преимущественно буквы, общие для русского и латинского алфавитов и понятные всем. Таких букв имеется 10-А, В, С, Е, Н, К, М, Р, Т, X. При употреблении только русских или только латинских букв в письменных сообщениях очень часто возникают недоразумения и путаница.
Кроме номера, на метках обычно ставят еще инициалы (буквы) того учреждения, которое производит работы. Если метят в открытых водоемах и меченые экземпляры могут быть пойманы иностранными судами или рыбаками, то на метке обязательно должно быть обозначено сокращенное название страны, в которой производят мечение. Советский Союз обозначают «USSR» или за недостатком места – одной буквой «R».
Скобочная метка изображена на рис. 78. Эту метку делают из монель-металла, никеля или посеребренной (луженой) латуни. Она представляет собой изогнутую пластинку с острым язычком на одном конце и с отверстием для него на другом.
Для большинства более крупных объектов метки делают длиной (в развернутом виде) 45 мм и шириной 7 мм и примерно 30 мм длиной и 5 мм шириной для более мелких (вобла и т. и.). Па метки для крупных объектов употребляют латунь толщиной 0,5-0,7 мм, а для мелких-0,4-0,5 мм.
Чем легче метки, тем лучше, поэтому толщину пластинки следует брать такую, чтобы язычок метки легко прокалывая ту часть тела, которую предполагают метить; но толще 0,7 мм делать метки не рекомендуется, так как они будут тяжелы. Номер на этой метке обычно ставят на одной половине, а инициалы на другой. При мечении в открытых водоемах инициалы удобнее помещать сверху (на половине с язычком), а во внутренних водоемах внизу (на половине с отверстием).
При заказе меток необходимо требовать, чтобы направление язычка строго соответствовало отверстию на другом конце, так как иначе метки не будут запираться (серебрят и лудят уже готовые метки).
Метки укрепляют на рыбе особыми щипцами, один из типов которых изображен на рис. 78.
Такие щипцы делают из обыкновенных плоскогубцев с подходящими по длине и ширине губками. На одной из губок (на рисунке-нижней) делают выемку с небольшими бортами и упором у заднего края.
![]() Рис. 78. Скобочная метка и щипцы для нее
Рис. 78. Скобочная метка и щипцы для нее
(по Караваеву, 1958):
а – общий вид щипцов со вставленной между губками меткой; б – внутренняя сторона нижней губки щипцов с выемкой; в– вид еще не согнутой метки; г – вид метки снизу; е – вид сверху.
У переднего края этой губки делают небольшое углубление, сходящее на нет к заднему краю губки. Когда метку сжимают щипцами, язычок входит в отверстие на другом конце метки и, скользя по углублению, загибается и запирает метку. В другой губке (на рисунке – верхней) делают большую выемку, чтобы при закрытии щипцов загибался только язычок, а сама метка не повреждалась и чтобы задний упор свободно проходил на нижней губке. В передний край этой выемки метка упирается сгибом у язычка. Благодаря этим двум упорам и бортикам на нижней губке метка прочно держится в щипцах при их слабом сжимании.
Чтобы избежать случайных ранений рыбы, края губок у щипцов должны быть тщательно закруглены, особенно у нижней губки, которую при мечении жаберной крышки вводят под жаберную крышку.
Во время работы (особенно в лодке, на плавучем садке и т. д.) щипцы из предосторожности следует привязывать к руке или поясу шнуром, продетым в отверстие, сделанное в одной из ручек щипцов.
Скобочную метку прикрепляют или к жаберной крышке или к хвостовому плавнику (последнее не рекомендуется).
При мечении в хвостовую часть метку сажают в основание лучей верхней лопасти хвостового плавника так, чтобы она захватывала и мускулы хвоста. Метка, посаженная на самый плавник, может легко сползти с его лучей. При мечении небольших рыб надо следить, чтобы метка не задевала последних хвостовых позвонков.
На жаберную крышку метку прикрепляют или на середине заднего края крышки или ближе к верхнему концу жаберной щели. Метка должна захватывать крышку на всю свою длину, но не очень сильно надавливать на край крышки. Во время мечения следует обращать внимание на положение жабр, чтобы не прихватить их меткой.
У некоторых рыб (например, сазана) кости жаберной крышки очень прочны, так что язычок метки из достаточно толстой пластинки не может ее проколоть. В таком случае следует сперва просверлить в жаберной крышке небольшое отверстие коротким шилом или кончиком ножа, можно даже не делать сквозного отверстия, а лишь наметить его двумя-тремя оборотами граненого шила или ножа, после этого язычок метки уже свободно прокалывает кость.
После наложения метки необходимо удостовериться, что она заперлась правильно, и если этого не произошло, то, сняв и выправив метку, надо пометить рыбу вновь.
Для того чтобы метка была лучше заметна на рыбе, н ней рекомендуется добавлять яркий цветной диск толщиной 0,2- 0,3 мм из целлулоида или пластмассы диаметром 10-15 мм (в зависимости от размера метки и объекта мечения). Этот диск (с небольшим отверстием в середине) перед наложением метки надевают туго на язычок метки. Язычком метка удерживается на рыбе во время мечения. Вместо диска с той же целью можно добавлять небольшую полоску из тонкой эластичной цветной пластмассы (например, полиэтилена).
Челюстная метка-это собственно та же скобочная метка, которая укреплена на нижней челюстной кости. (Челюстная метка на рыбе, казалось бы, не имеет преимущества перед обычной скобочной меткой, однако по экспериментам в США с радужной и ручьевой форелью и другими речными рыбами эта метка оказалась более заметной, так как давала до 50% возврата). Ею можно метить те виды, которые обладают более прочной челюстной костью.
При укреплении метки скальпелем делают небольшой разрез снизу, с внутренней стороны нижней челюсти рыбы и в этот разрез вставляют конец (без язычка), а затем ее запирают обычным образом круглогубцами соответствующего размера (специальные щипцы в данном случае не требуются).
Для того чтобы метка не препятствовала росту челюстной кости (что может привести к разжатию и потере метки), вытянутый овал, получающийся после закрытия метки, расширяют почти до формы круга, нажимая круглогубцами .(или плоскогубцами) на оба конца овала.
Эту метку можно сделать из более тонкого металла, чтобы она была легче, лишь бы язычок был достаточно твердым и надежно запирал метку.
Метка Петерсена (для «плоских» рыб-камбалы и ската) получившая очень широкое распространение, состоит из двух эбонитовых или целлулоидных дисков, соединенных между собой серебряной или нержавеющей проволокой толщиной 0,7- 1,0 мм (рис. 79). На одном из дисков (верхнем) ставят номер и инициалы.
![]()

Рис. 79. Метка Петерсена (по Караваеву, 1958).
Рис. 80. Игла Ганзена со вставленной проволокой и Диском (по Караваеву, 1958).
Диски обычно имеют диаметр 15 мм, но диаметр может быть любого размера, в зависимости от величины объектов, подлежащих мечению. Края дисков должны быть ровные и гладкие (лучше закругленные), толщина дисков обычно 2- 3 мм, верхний диск может быть несколько тоньше.
У камбал эти метки укрепляют под основанием спинного плавника, в задней трети тела, причем проволокой прокалывают мускулы, а диски помещают один (с номером и инициалами) с верхней, «зрячей», стороны тела, а другой – с нижней, «слепой».
У скатов такие метки укрепляют в передней части тела в области грудных плавников.
Для закрепления метки концы проволоки с той и другой стороны загибают круглогубцами в небольшую петельку так, чтобы самый кончик был прижат петелькой к диску. Этот прием до некоторой степени гарантирует от соскакивания диска, даже если петелька почему-либо отогнется.
При укреплении метки мускулы прокалывают или самой проволокой, для чего ее обрезают кусачками так, чтобы кончик ее был достаточно острым (обрез идет косо), или же для этого применяют специальную иглу Ганзена (рис. 80), которую, подобно колпачку, надевают на конец проволоки и прокалывают вместе с ней. Эту иглу делают длиной 6-7 см и диаметром 2,5-3,5 мм. С тупого конца иглу просверливают вдоль на 4-5 см., причем диаметр канала должен быть немного больше диаметра употребляемой проволоки, чтобы проволока входила в канал совершенно свободно. Иглу делают из металла, не поддающегося действию морской воды.
Мускулы можно также прокалывать иглой от шприца для инъекций, канал иглы должен соответствовать диаметру проволоки. В этом случае, после того как мускулы проколоты иглой, проволоку вставляют в отверстие канала с острого конца иглы. Вынимая иглу обратно, на ее месте оставляют проволоку. Всегда должен быть достаточный запас этих игл. Чтобы избежать порчи и ржавления игл, по окончании работы их следует хорошо промыть пресной водой, смазать жиром. Если игла сильно затупится, ее можно точить на мелкозернистом бруске или в крайнем случае на наждачной бумаге.

Рис. 81. Метка для осетровых (по Караваеву, 1958).
Перед работой диски с номерами подбирают по порядку номеров в ящики или нанизывают на проволоку, нижние диски могут находиться в любой удобной таре вместе с кусочками проволоки необходимой длины. Один конец проволоки загибают петелькой, и она может быть заранее вставлена в нижний диск (чтобы проволока не выпала из диска, свободный конец ее отгибают в сторону).
Меткой Петерсена можно метить не только «плоских», но и «круглых» рыб. У круглых рыб эту метку обычно укрепляют у основания спинных плавников, причем пластинки помещают с той и другой стороны тела или же на жаберную крышку. В этом случае толщина пластмассовых дисков может быть 0,5-1,0 мм.
Кроме того, оба диска или один из них с номером могут быть изготовлены из металла толщиной 0,20-0,25 мм; в некоторых странах один диск (без номера) делают из раковин соответствующей толщины.
Некоторые исследователи для укрепления этой метки на жаберной крышке предложили специальные щипцы, но ни щипцы, ни сам этот способ не получили широкого распространения.
Метка для осетровых рыб и некоторых других представляет собой овальную пластинку (длина 25-30 мм, ширина 10- 15 мм) с двумя отверстиями на узких концах (рис. 81). Ее делают из цветного целлулоида, пластмассы или металла и укрепляют с помощью подходящей неокисляющейся проволоки.
У осетровых рыб эту метку укрепляют на спине между 2-й и 3-й и или 3-й и 4-й жучками, а у севрюги, кроме того, она может быть укреплена на верхней стороне рыла, несколько ближе к его кончу (впереди линии усиков).
При укреплении метки проволоку вводят в отверстие, которое делают обычным шилом или шилом с отверстием у острого его конца (подобно игле для швейной машины), иглой Ганзена, или большой иглой для инъекций.
В некоторых случаях этой меткой можно пользоваться для мечения других рыб, особенно когда оно рассчитано на относительно короткий срок (1-2 месяца), например для ходовых лососей. В этих условиях овальную пластинку укрепляют относительно мягкой проволокой просто на хвостовом стебле. На короткий срок можно использовать «эмалированную» проволоку из красной меди (марка-«эмалированный провод обмоточный») .
Метка для угря – небольшая овальная пластинка, с одной стороны вытянутая в длинную ножку или плоскую иглу (рис. 82). Примерный размер пластинки:
длина 13-15 мм, ширина около 7 мм, длина ножки 17-20 мм. Метки делают из серебра или одного из указанных выше металлов; на пластинке выбивают номер и инициалы.
![]()
Рис. 82. Метка для угря (по Караваеву, 1958).
Метку укрепляют в коже вдоль основания спинного плавника, предварительно делают два прокола кривым плоским шилом, так как кожа угря очень плотная. Метку вставляют в прокол, расположенный ближе к голове, тонкий конец метки (ножку) выводят наружу через второй прокол и соединяют пластинку с ножкой.
Во время мечения угря кладут на сухое полотенце или пропускную бумагу, на которых он лежит совершенно спокойно и не мешает работе.
Подвесные метки довольно разнообразны по форме, размеру и материалу, но в большинстве случаев имеют вид продолговатой овальной или каплевидной пластинки (рис. ,83).
Эти метки прикрепляют, как правило, в области спинного плавника, впереди или сзади него, или между спинными плавниками нержавеющей проволокой толщиной 0,2-0,5 мм, или капроновой (нейлоновой) ниткой, или жилкой примерно такой же толщины.
Подвесные метки делают из тонкого листового металла толщиной примерно 0,2 мм (серебро, монель-металл, нержавеющая сталь и др.), целлулоида или другой стойкой к морской воде пластмассы (толщиной 0,3-0,6 мм), а также из плотной бумаги или нетолстого картона, покрытых в готовом виде целлулоидным лаком (Целлулоидный лак можно получить, растворив в ацетоне кино- или фотопленку, предварительно отмытую от фотоэмульсии) или нитролаком.
Размер подвесных меток: длина 1,3-3,0 см, ширина 0,4- 0,6 см, а каплевидных – до 1,0 см в широкой части.
При укреплении меток проволоку или жилку проводят через мускулы у крупных рыб шилом с отверстием, у более мелких – при помощи иглы для инъекций или хирургической иглы.
Рис. 83. Подвесные метки (по Караваеву, 1958):
а-малая овальная; б-продолговатая пластмассовая пластинка; в-каплевидная пластмассовая; г-бумажная на фотокартоне; д — из тонкой целлулоидной пластинки.
На металлические подвесные метки набивают номера и инициалы, обозначающие название страны или исследовательского учреждения. На целлулоидных метках номера и инициалы выдавливают штампом. В этом случае нагревают штамп, или осторожно разогревают целлулоид. Можно их писать тушью от руки, а затем покрывать целлулоидным лаком (нитролаком), или инициалы страны могут быть выдавлены при изготовлении (штамповке) самих меток, а тушью пишут только порядковые номера. (Если надписи тушью сделаны только с одной стороны пластинки, то метку покрывать надо с обеих сторон, так как иначе высыхающий лак погнет ее в одну сторону, особенно если пластинка тонкая.)
Бумажную подвесную метку (рис. 84, г) делают из толстой фотобумаги (фотокартон). В этом случае на одной стороне помещают сильно уменьшенный краткий фототекст с названием и адресом учреждения, куда следует послать метку с необходимыми сведениями, и сообщается о выплате премии за каждую доставленную метку. Порядковый номер пишут тушью на другой стороне метки.
Для некоторых рыб подвесные метки можно изготовлять из тонкого целлулоида, имеющего меньший вес, но несколько больший размер, примерно 3,3х1,5 см (рис. 83, д).

Рис. 84. Гидростатическая метка Э. Леа (по Караваеву, 1958).
На прозрачную целлулоидную пластинку метки с той и другой стороны наклеивают (нитролаком или целлулоидным лаком) полоски бумаги с напечатанным на них кратким текстом и на одной из полосок пишут порядковый номер тушью.
Чтобы эта метка была лучше заметна на рыбе, бумагу следует употреблять ярко-желтого цвета, а свободные части метки выкрасить в ярко-синий или голубой цвет. У одного края метки делают отверстие для проволоки или капроновой жилки. В случае прикрепления метки проволокой, чтобы она не прорывала тонкую пластинку, следует подклеить небольшие кусочки такого же целлулоида между самым краем метки и отверстием с одной или обеих сторон. Готовую метку следует покрыть лаком с обеих сторон.
Гидростатическая метка Э. Леа – та же подвесная метка, но более совершенная. Благодаря помещаемой внутри подробной записке-инструкции, как поступить с найденной меткой, обеспечивается наибольший возврат меток-до 20-25% (рис. 84).
Эта метка состоит из желтой прозрачной целлулоидной трубки диаметром около 4 мм, в которую вложена свернутая записка. Концы трубки тщательно заделаны и окрашены нитрокраской в синий или голубой цвет. На одном конце имеется петелька, за которую проволокой или жилкой метку укрепляют на спине рыбы в области спинного плавника. Проволоку или жилку проводят через мускулы, как указано выше.
Длина метки в готовом виде 30-50 мм. В запечатанной трубке содержится некоторое количество воздуха, поэтому метка плавает и называется гидростатической.
Через прозрачный целлулоид должен быть виден край скрученной записки, на которой напечатаны 3-4 строчки на 2- 3 языках: «обрежьте края, письмо внутри», а также условные буквы страны и порядковый номер. Этот номер и такой же номер на средней части записки, пишут от руки тушью.
Рис. 85, Внутренние метки: а – металлическая; б-целлулоидная (по Караваеву, 1958).
В основной части записки указано, куда и кому послать найденную метку и какие сведения необходимо сообщить: место, время, глубину вылова, орудие лова, которым поймана рыба, ее размер, указание о том, как измерить рыбу, просьба приложить отолиты или чешую пойманной рыбы, а также указать свое имя и адрес. В записке также указывается, что за присылку метки и сообщенные данные выплачивают премию.
В тех случаях, когда эту метку применяют в районе международного рыболовства, широко освоенного многими странами, записку следует печатать на 2-4 языках. Текст должен быть кратким и понятным.
Для того чтобы на рыбе были лучше заметны метки (особенно при больших уловах), к некоторым видам меток можно добавлять небольшие полоски («флаг») из тонкой эластичной пластмассы яркого цвета, например, хлорвинила. Лучше всего для этого употреблять полоски ярко-оранжевые или ярко-желтые и в некоторых случаях голубые или красного цвета, в зависимости от вида рыбы, условий промыслового лова и других сопутствующих обстоятельств.
Полоски для крупных видов могут быть примерно длиной до 10 см и шириной 1-2 см. Эти полоски можно добавлять к гидростатическим, петерсеновским (при укреплении их на жаберной крышке или на спине), а возможно и к скобочным и другим меткам.
Внутренняя (брюшная) метка представляет собой небольшую металлическую пластинку с закругленными концами, которую через небольшой разрез в боку вводят в брюшную полость рыбы. На ней выбит порядковый номер и условные буквы страны (рис.85).
Металлические внутренние метки (никель, никелированная сталь или нержавеющая сталь некоторых марок, имеющая магнитное свойство) длиной 10-20 мм, шириной 3,0-4,5 мм и толщиной 0,5-1,5 мм употребляются во многих странах преимущественно для сельди и сардины, а также некоторых других видов рыб, которых перерабатывают на специальных заводах на жир и кормовую муку.
Металлические внутренние метки обнаруживают и собирают при переработке рыбы на утилизационных заводах электромагнитами или специальными детекторными установками, которые установлены на транспортерах предприятий.
У нас рыбу на утилизационных заводах не перерабатывают, поэтому мечение внутренними металлическими метками не применяют.
Внутренними метками более крупными, примерно длиной 4,0-6,0 см, шириной около 1 см и толщиной около 0,5 см, изготовленными из целлулоида и ярко окрашенными, иногда метят более крупных рыб (рис. 86, б), в частности треску и пикшу, которых в основной массе обрабатывают непосредственно на рыболовных судах, и метки могут быть обнаружены при удалении внутренностей (шкерка). В соответствующих условиях этими метками можно метить и других промысловых крупных рыб.
Относительно большие размеры этой метки позволяют на ней обозначить не только обязательный порядковый номер и условные буквы страны, но также название исследовательского учреждения, его адрес, сообщение о выплате премии за доставку метки и другие сведения.
Серийное мечение
В тех случаях, когда мечение рассчитано на сравнительно короткий срок и его задачей является выяснение какого-нибудь ограниченного вопроса, например определения скорости подъема по реке проходных рыб, ската их после нереста, распределение стада на нерестилищах и т. д., нет необходимости употреблять нумерованные метки, можно обойтись более простым способом мечения.
В таких случаях целую партию рыбы в каком-нибудь ограниченном районе и в течение короткого срока метят одинаковой меткой.
Для серийного мечения можно употреблять подходящей толщины нержавеющую проволоку или цветные капроновые жилки (или нитки). Метки укрепляют в спине под основанием спинного плавника (в передней или задней его части). Концы прошитой через мускулы проволоки или жилки скручивают или завязывают узлом со стороны спины. То и другое прошивают хирургической медицинской иглой или обычной с ушком, а проволоку, кроме того, можно продеть иглой Ганзена у крупных рыб или иглой для инъекций у более мелких.
Если необходимо различать несколько партий, помечаемых одновременно в различных пунктах или разновременно в одном и том же пункте, то жилки или нитки употребляют разных цветов. При большом числе серий можно метить сразу двумя-тремя жилками, комбинируя различные цвета.
При мечении проволокой с этой же целью употребляют разноцветные стеклянные (литые) или пластмассовые бусинки, нанизывая их на проволоку. При большом числе серий также комбинируют цвета бусинок, надевая их на проволоку по одной – три штуки.
Мечение молоди
На сравнительно ранних стадиях (соответствующих стадии парр) преимущественно молодь лососей метят срезанием плавников или их частей острыми тонкими ножницами или маникюрными кусачками для ногтей, при этом обязательно надо удалять и основные косточки плавниковых лучей, так как иначе лучи могут отрасти. (‘ Практиковавшееся иногда мечение молоди путем срезания части жаберная крышки не достигает цели, так как жаберная крышка сравнительно легко восстанавливается.).
Такое мечение надо проводить, возможно, быстрее, весьма аккуратно и осторожно, чтобы избежать большой гибели помеченных мальков. Во время самой операции срезания плавников малька держат в мокрой вате.
Лов мальков перед мечением также должен быть организован с максимальными предосторожностями.
Для того чтобы избежать ошибок при вылове уже взрослой рыбы и не принять от природы уродливых особей за меченых, следует срезать одновременно два каких-нибудь плавника, например часть спинного и жировой или часть спинного и один брюшной.
Если молодь метят из года в год или одновременно в различных пунктах, то для того, чтобы отдельные партии можно было различить, плавники срезают в разных комбинациях. Спинной и анальный плавники обычно срезают только наполовину, удаляя их переднюю или заднюю часть. Отрезая и удаляя один из брюшных, а для лососевых еще и жировой, можно получить достаточное количество комбинаций.
Если метят более взрослую молодь (соответствующую стадии смолт), то используют нержавеющую проволоку (одну или с бусинками), вводя ее под основание спинного плавника, как для серийного мечения. Толщину проволоки, и размер бусинок подбирают в соответствии с величиной объектов мечения.
Подросшую молодь рациональнее метить подвесными метками с порядковыми номерами (целлулоидными, металлическими или бумажными). Эти метки имеют длину 1,3-2,0 см и ширину 0,4-0,5 см (рис. 86).
С постепенным ростом особи, помеченной таким образом, проволока обычно все время остается поверх спины, а часть, проходящая сквозь тело, поднимается кверху, прорезая мускулы и оставляя по бокам темный след (шрам), по которому, в случае потери проволоки или метки, легко обнаружить, что рыба носила метку.
Спинной плавник Узел проволоки
 метка
метка
Проволока
Рис. 86. Прикрепление метки к молоди
семги (по Мельниковой).
При мечении молоди подвесными метками можно также прикреплять их, прокалывая мускулы спины двумя концами проволоки, при этом сама метка остается на одной стороне, а концы проволоки закручивают с другой стороны спины. Оба конца проволоки одновременно пропускают через мускулы при помощи двух игл для инъекции, спаянных вместе на нужном расстоянии.
М. Н. Мельникова (ГосНИОРХ) для мечения молоди семги использовала метки из целлулоида желтого цвета толщиной 0,5 мм и размером 4х11 мм. Надписи на метках делались тушью: на одной стороне номер, на другой сокращенное название учреждения. Затем метки покрывались раствором фотографической пленки в ацетоне.
Для прикрепления меток употреблялась нержавеющая проволока толщиной 0,3-0,4 мм (пригодна спираль электроплитки). Концы проволоки срезались косо и проволока проходила через ткани рыбок без помощи иглы.
Проволока загибалась в виде буквы U, на нее. надевалась метка и оба конца пропускали через мышцы под задней третью спинного плавника, с захватом его птеригиофоров (см. рис. 86).
Для того чтобы молодь легче переносила мечение, применялся наркоз уретаном: перед мечением рыбки помещались в таз с водой, к которой постепенно добавляли раствор уретана до концентрации примерно 2%. При мечении рыбку обертывали мокрой марлей. Метили вечером при более низкой температуре.
При мечении рыб, которые значительно вырастут (например, лососи), между самой меткой и двойной проволокой вставляют дополнительный кусочек проволоки, чтобы проволока, удерживающая метку, не сильно врезалась в мускулы.
Для мечения осетровой молоди можно применять вылущивание некоторых спинных или боковых жучек, но не меньше двух в ряду, чтобы в дальнейшем отличать меченые экземпляры от случайно потерявших жучки. Для того чтобы различать несколько партий меченой молоди (по районам или годам), следует твердо устанавливать, какие жучки в ряду вылущивать.
Для контроля выживания меченой молоди от каждой партии оставляют некоторое количество мальков и выдерживают их некоторое время. Это возможно, главным образом, при мечении мальков, искусственно выведенных на рыбоводных заводах.
Для обеспечения достаточного возврата необходимо метить одновременно значительное число молоди, т. е. несколько десятков тысяч, имея в виду ее большую естественную гибель. Следует также широко информировать всех лиц, имеющих отношение к рыболовству, так как эти метки менее заметны и легче могут быть пропущены при вторичной поимке, особенно несведущим лицом.
Обработка результатов течения
При обработке результатов мечения следует пользоваться особой карточкой, заполняемой на каждый вторично пойманный экземпляр. В эту карточку вносят все сведения, относящиеся к этому экземпляру в момент мечения и параллельно-в момент вторичной поимки (первые выписывают из журнала мечения, а вторые – из карточки, заполненной при сдаче метки).
Ниже помещаем примерный образец такой карточки. Эта карточка может быть дополнена еще пунктами о времени, протекшем между мечением и вторичной поимкой, пройденном расстоянии, среднесуточной скорости миграции, направлении миграции или другими сведениями в зависимости от целей исследования. Употребление таких карточек значительно облегчает и ускоряет работу.
При изучении миграций по результатам мечения сведения о вторичных поимках для большей наглядности наносят на карту. Обычно на ней отмечают место выпуска меченых рыб жирным кружком или квадратиком, а пункты поимки меченых экземпляров – небольшими кружками или точками. Или же результат мечения изображают пучком линий или стрелок, выходящих из пункта мечения и заканчивающихся там, где были пойманы меченые рыбы. Эти линии изображают предполагаемый или допускаемый путь миграции одной или группы рыб.
Если при таком изображении нужно как-нибудь выделить отдельные группы вторичных поимок или же разделить материал для сравнения по тем или иным признакам (например, отделить самцов от самок, разделить по зрелости, по длине, по сезонам лова, по способу вторичной поимки, по принадлежности той или иной стране судов, доставивших метки и пр.), то пункты вторичных поимок соответственно обозначаются разными значками или же, что нагляднее, различными цветами.
Образец карточки
|
Метка _____________________________ №._____________ Учреждение Название рыбы ________________________________________________ |
||
|
Наименование |
При мечении |
При вторичной поимке |
|
Название судна, рыбозавода Номер станции Место (координаты, номер квадрата) Дата Глубина, м Температура Соленость, %о Орудие лова Длина рыбы, см Вес рыбы, г Возраст Пол |
||
В некоторых случаях достаточно эти дополнительные сведения (правда, с ущербом для наглядности) отмечать на карте около мест вторичных поимок. Например, указать дату и месяц вторичной поимки этого экземпляра или поставить рядом цифру, обозначающую число месяцев или дней между мечением и вторичной поимкой, или же отметить буквой национальную принадлежность судна, поймавшего данный экземпляр и т. д.
Теперь начинают применять метод радиоактивных индикаторов, получивший наименование метода меченых атомов. Но для применения такого метода необходимы серьезные специальные знания о радиоактивных веществах (радиоактивных изотопах), полное знакомство со способами пользования этими веществами. Нужны соответствующие приборы и исключительные предосторожности со стороны занимающегося радиоактивными веществами. Методы мечения рыб радиоактивными веществами изложены А. С. Трошиным (1956).
Глава Х
УЧЕТ СОСТОЯНИЯ РЫБНЫХ ЗАПАСОВ И ОПРЕДЕЛЕНИЕ ВОЗМОЖНЫХ УЛОВОВ
Краткий обзор методов определения рыбных запасов и прогнозирования уловов
Методика определения запасов рыбы продолжает оставаться одной из самых трудных и наименее освещенных ихтиологических проблем, хотя попыток определения рыбных запасов делалось много и имеется большое количество работ, рассматривающих эту проблему.
О состоянии промысловых запасов рыб до известной степени можно судить по анализу возрастного состава. Присутствие в уловах младших возрастных групп указывает на неблагополучное состояние запасов, а старших возрастов – на недостаточное использование запасов. По данным К. И. Беляевой (1959) судака северной части Онежского озера добывали преимущественно в возрасте свыше 10 лет, такой судак составлял до 69% общего вылова этого вида рыбы. Таким образом, можно смело говорить о недолове судака, о том, что запасы его позволяют повышать уловы.
Подобные выводы возможны лишь в тех случаях, когда исследователь имеет материал, полученный из уловов такими орудиями, которые могут брать рыб различных размеров (различных возрастов), начиная с мелких, неполовозрелых особей и
кончая самыми крупными.
Наиболее ценен материал неводного лова и материал, обширный в количественном отношении. Нужно установить возрастные группы в уловах, если не каждого месяца, то, по крайней мере, по периодам года (по путинам), чтобы дать средние показатели возрастных группировок за целый промысловый год.
Е. В. Бурмакин и Г. В. Домбровский (1956), проанализировав многолетние данные по рыбам Балхаша (возрастной состав, размеры и вес, количество повторно созревающих и повторно нерестующих рыб), приходят к выводу, что «размерный и возрастной состав основных промысловых рыб озера Балхаша свидетельствует о длительном недоиспользования промысловых возможностей», и рекомендуют усиливать промысел.
Приведенные соображения могут в общей форме определить состояние рыбных запасов, но количественные показатели, выводимые из подобных соображений, математической достоверности под собой не имеют. Устанавливать так называемые возможные уловы, выражая их в числовых показателях (в тоннах или центнерах), только на основании констатирования недолова или перелова крайне рисковано. Однако даже общие суждения о запасах имеют большое значение.
Н. М. Книпович (1938) говорит: «производя из года в год анализы возрастного состава известного вида рыб в определенном районе, возможно, бывает совершенно ясно проследить на протяжении ряда лет выраженное влияние обильного или, напротив, скудного приплода в определенный год». Урожайные или неурожайные годы могут встречаться у разных видов рыб, и они, естественно, влияют на состояние запасов, и это влияние необходимо учитывать при суждении о состоянии запасов по возрастной группировке рыб в промысловых уловах и пытаться найти причины колебаний запасов рыб.
После того как в водоеме хотя бы в общей форме дана оценка состояния запасов рыб ихтиолог может заниматься целым рядом биологических и промысловых вопросов, имеющих непосредственную связь с рыбными запасами. Если доказано, что запасы состоят преимущественно из рыб младших возрастов, или только из старших, – нужно разработать мероприятия по накоплению отсутствующих возрастных групп рыб. В этом отношении будут полезны предложения покровительственного характера вплоть до установления запретных сроков и мест для лова того или другого вида рыб. Если водоем заполнен малоценной сорной рыбой, – нужно разработать меры для снижения или, где это возможно, для полного подавления запасов сорных рыб. Для этого предлагают биологические методы (введение в водоем хищных рыб или покровительство имеющимся в водоеме хищникам), методы технические (усиленный вылов сорной рыбы), химические или, наконец, уничтожение выметанной икры ненужных рыб. К такого рода мероприятиям следует отнести охрану нерестилищ и разведение более ценных рыб.
Состояние запасов любого вида рыб в сильной степени зависит от биологических свойств данного вида и от его взаимоотношений с другими рыбами и с беспозвоночными, населяющими тот же водоем.
Рыбы, у которых половая зрелость наступает в раннем возрасте, обычно имеют большую численность, и промысловые запасы их велики (тихоокеанская горбуша, ерш).
Поэтому при изучении состояния запасов рыб необходимо устанавливать время наступления половой зрелости каждого вида рыб, обитающего в исследуемом водоеме, помня, что сроки наступления половой зрелости у одних и тех же видов в различных водоемах различны (Ефимова, 1954). Например, лещ в южных водоемах созревает в возрасте 3-4 лет, а лещ в водоемах Карелии, как установила на очень большом материале О. И. Потапова (1954), половозрелости достигает на 8-9-м году жизни. Причем этот же исследователь доказал, что карельский лещ сохраняет воспроизводительную способность до своего предельного возраста, каким Потапова называет 26 лет.
Такие вопросы биологии рыб, как время наступления половой зрелости, плодовитость индивидуальная и видовая, продолжительность сохранения воспроизводительной способности, естественная смертность должны служить существеннейшей частью исследований состояния запасов рыб.
Межвидовые взаимоотношения рыб относятся к тем биологическим факторам, которые в очень большой степени влияют на состояние рыбных запасов. Хищные рыбы истребляют рыб мирных: отложенную и развивающуюся в водоеме икру, вышедших из икры личинок, мальков и, наконец, взрослых рыб во все периоды их жизни. Нет оснований исключать и внутривидовую конкуренцию рыб: сиги поедают сами ими отложенную икру, это явление, несомненно, свойственно и другим видам рыб, щуки поедают рыб своего же вида. Такая форма внутривидовых отношений, правда, не столь сильно сказывается на запасах рыб, как формы межвидовой конкуренции. Исследовать межвидовые взаимоотношения нужно в природе, в самом водоеме, а не в аквариумах, так как в аквариумах совершенно искажается истинная картина взаимоотношений рыб.
Изучение межвидовых взаимоотношений требует сбора материалов по питанию хищничающих рыб за круглый год и по всем возрастным их группам, с определением количеств потребляемой ими пищи (в виде икры, молоди и взрослых рыб) за сутки, месяц, за год. Очень важно выяснить связь прироста веса хищников с количеством потребляемого ими корма. Интересные исследования межвидовых взаимоотношений озерных рыб проводились ихтиологами и гидробиологами Института биологии Карельского филиала Академии наук СССР (Бала-гурова,1963).
На состояние запасов рыб большое влияние оказывает приплод молоди. Хорошие урожаи молоди способны повысить запасы взрослых рыб. Учет молоди рыб (на примере молоди воблы) В. С. Танасийчук (1940) советует вести путем сравнения средних уловов молоди за 1 ч траления стандартным мальковым тралом.
Автор при составлении прогноза запасов промысловых рыб использует многолетние данные по количественному учету урожая молоди и показывает, что между урожаем молоди и мощностью поколений у рыб есть зависимость, но эта зависимость непостоянна вследствие непостоянства кормовых ресурсов. На основании этого автор приходит к выводу, что величина стада рыб зависит не только от количеств отложенной икры и вышедшей из нее молоди, но и от условий нагула молоди, пастбищ.
О процессе колоссальной гибели молоди рыб в естественных условиях можно судить по таким строкам, приведенным В. В. Васнецовым (1946): «Из выметанной лещом икры в количестве 100000 штук на третий день погибло 13%. Через 20 дней после выклева осталось только 10000 личинок, т. е. гибель за это время равнялась 90%, а через два с половиной месяца погибло 99,7%».
Одним из важнейших факторов, влияющих на состояние рыбных запасов, признается количество кормовой биомассы. Поэтому при всех ихтиологических работах надо определять количество бентоса в водоеме. Но продуктивность бентоса в водоеме нередко определяется на основании краткосрочных гидробиологических работ, вследствие чего не отражают действительности. Требуются наблюдения круглогодичные и в продолжении нескольких лет. Таких работ пока очень мало. Для примера можно привести работу Е. В. Боруцкого (1946). Исследования велись систематически три года на Белом Косинском озере. Автор показал, что лучшим показателем продуктивности и кормности водоема может служить весенняя биомасса бентоса, кроме того, он указал метод прогнозирования состояния бентоса весной следующего года. «Для этого необходимо иметь следующие данные за предшествующий год: 1) весеннюю биомассу, полученную на основании дночерпательных серий ранней весной перед вскрытием озера, 2) наличие или отсутствие весеннего перемешивания воды в водоеме, 3) термическую стратификацию в августе для определения степени прогрева придонных слоев водоема, 4) температурные условия и продолжительность осеннего периода».
Ихтиологу, занимающемуся запасами рыб, необходимо знать, насколько тот или другой вид рыб обеспечен кормом. Н. И. Чугуновой было установлено, что запасы бычков в Северном Каспии в 1941 г. по сравнению с 1934- 1935 гг. уменьшились. Выясняя причины этого уменьшения, автор остановил свое внимание и на повышении солености воды в рассматриваемом участке моря (в восточных частях Северного Каспия) и на уменьшении бентоса, которым бычки питаются, (уменьшение произошло, по-видимому, вследствие осоленения, и, может быть, вследствие замора из-за снижения содержания кислорода). Автор приходит к выводу, что отрицательное влияние на численность бычков оказало уменьшение бентоса, уменьшение пищи бычков. «Уменьшение количества пищи, очевидно, вызывает усиление конкуренции в пище; рыбы не докармливаются, что должно отрицательно действовать на их рост, скорость g, созревания, плодовитость, выносливость молоди и т. п., а в конечном счете, и на их численность» (Чугунова, 1946).
Не меньшее значение в питании многих промысловых рыб имеет и планктон, который в начальные периоды жизни молоди рыб (личинок и сеголетков) служит основным кормом.
Гидрологические условия б водоеме непостоянны, и изменения их влияют на состояние рыбных запасов. Н. М. Книпович (1938) справедливо говорил: «промысловая продуктивность водоема определяется совокупностью гидрологических и тесно связанных с ними биологических условий, как и общая продуктивность его».
Деятельность человека, выражающаяся в изъятии рыб из водоема, оказывает глубочайшее влияние на видовой состав рыбного стада, на количество и качество отдельных видов.
Изучение состояния рыбного стада в водоемах, которые не используются промыслом, даст ценнейшие основания для регулирования этого стада в период введения промысла. Однако о состоянии запасов рыб приходится судить обычно тогда, когда в водоеме естественные соотношения возрастных и видовых группировок достаточно нарушены промыслом. Но все же есть примеры, когда ихтиолог располагает и допромысловыми сведениями. Такой пример можно видеть в работе П. А. Моисеева (1946). Пример заслуживает внимания с методической стороны. Изучение состояния стада камбал залива Петра Великого началось одновременно с организацией лова, т. е. с 1929 г. и продолжалось до 1946 г. – более 15 лет. Наблюдения велись за местами лова, за видовым и количественным составом уловов, за возрастными, линейными и весовыми группировками камбал в уловах, рыб метили, изучали их питание и плодовитость. Через 15 лет работы большими траулерами и маломощными судами при помощи снюрревода оказалось, что произошли существенные изменения в размерах, составе, поведении и биологии камбал, сильно изменились уловы и в своих общих показателях, и в средних за сутки и за час работы орудия лова. Все это было подмечено в первые же пять лет работы промысла. Площади зимних скоплений камбал стали меньше, что подтверждает уменьшение стада. Численность камбал старших возрастов уменьшилась и соответственно увеличилось количество рыб младших возрастов. Вместе с тем установлено, что темп роста камбал повысился, что ставится в связь с уменьшением запасов камбал, так как оставшиеся особи освободились от напряженной пищевой конкуренции. Главный вывод из своих наблюдений автор формулирует такими словами: «Следует стремиться создать такое положение, чтобы для любого вновь осваиваемого или уже давно облавливаемого стада рыб был достигнут режим, заключающийся в первую очередь в установлении ежегодного размера вылова при сохранении нормального соотношения возрастных групп стада. Ни в коем случае нельзя допускать перелова, следствием которого является исчезновение старших возрастов и перенос центра тяжести на молодые возрастные группы, что сводит на нет положительные стороны некоторого разрежения стада».
Приведенные примеры показывают насколько необходимы одновременные и продолжительные наблюдения ихтиологические, гидробиологические, гидрологические и промысловые, когда нужно заниматься вопросами о состоянии запасов промысловых рыб и об установлении норм вылова. Однако повторяем, – современное состояние ихтиологической методики еще не позволяет говорить о точных числовых показателях запасов рыб, но судить о состоянии этих запасов уже стало возможным.
П. А. Дрягин (1951) советует пользоваться при определении рыбных запасов следующими признаками состояния биологического процветания или угнетения того или другого вида данного водоема.
|
Основные признаки биологическогопроцветания вида |
Признаки угнетения |
|
Средняя навеска в уловах высокая |
Средняя навеска снижается. |
|
Темп роста нормальный, постепенно снижающийся. |
Темп роста замедленный, либо обратный нормальному, либо резко колеблющийся. |
|
Высокая упитанность и хороший экстерьер в период нагула. |
Понижение упитанности, ухудшение экстерьера. |
|
Доминирует над конкурентами. |
Подавляется конкурентами |
|
Количество самок в меньше количества самцов. |
Преобладают самцы. |
|
Высокая промысловая численность, сравнительно мало изменяющаяся по годам. |
Сокращение промысловой численности, частые ее колебания. |
|
У некоторых видов наблюдается распад на локальные стада. |
Происходит сокращение и иногда исчезновение отдельных локальных стад. |
Решающее значение для методики определения рыбных запасов и для прогнозов уловов имеет многолетняя статистика уловов. Значение такой статистики было бы еще больше, если бы уловы приводились не только в весовых, но и в штучных показателях: тогда легче было бы судить о численности- промыслового стада рыб. Примером поштучного учета рыб служат показатели вылова дальневосточных лососей.
В настоящее время особенно остро ставятся вопросы, связанные с состоянием запасов рыб, ибо запасы лимитируют уловы, а уловами определяется и весь характер рыбного хозяйства того или другого водоема.
Г. В. Никольский (1950 а) сделал, краткую сводку работ, вернее направлений, по проблеме динамики численности стад и изложил свой взгляд, причем, основные положения предлагаемой им трактовки являются в значительной степени коллективным трудом сотрудников кафедры ихтиологии Московского Государственного университета. С методической стороны статья Г. В. Никольского для ихтиологов крайне поучительна. Для суждения о состоянии стада какого-либо вида рыб, а также для определения возможной величины вылова прежде всего необходимо знать «наибольший темп роста, который может дать .вид в данном водоеме, и наиболее ранний возраст половозрелости, которого вид может достигать».
«Для лососевых, карповых, осетровых, камбаловых и, видимо, ряда других групп рыб установление контингента должно базироваться на следующих показателях: 1) темп роста должен быть близок к максимальному видовому и лишь немного меняться из года в год; 2) время наступления половозрелости должно быть у большинства особей наиболее раннее, свойственное данному виду; возрастной ряд впервые созревающих рыб должен быть максимально сжатым; 3) кормовая база (которая учитывается перед началом откорма) должна быть сходна с предыдущими годами; 4) уловы из года в год остаются относительно стабильными при постоянном (не уменьшающемся) количестве орудий лова».
Далее автор поясняет: «Если все эти показатели имеются налицо, то улов может быть дан равным прошлогоднему. Если же темп роста высокий, промысел базируется на первых половозрелых группах, половозрелость ранняя, кормовая база высокая, и уловы падают-необходимо значительное снижение уловов и улучшение условий воспроизводства с последующей проверкой биологических показателей учитываемого вида, состояния кормовой базы и изменения численности других (особенно не промысловых) видов, питающихся сходной пищей с видом, запасы которого подорваны. При замедлении темпа роста, поздней половозрелости, растянутости возрастного ряда впервые созревающих особей, значительном выедании кормовых объектов, стабильности уловов и увеличении уловов на одно орудие-возможно увеличение величины вылова».
При определении контингента вылова советуется брать одновременно группу видов, «связанную биологией размножения и пищевыми взаимоотношениями».
Пока нет инструкций, в которых были бы изложены все нужные методические указания по определению и прогнозированию запасов и уловов рыб. Наиболее существенными работами, которые могут быть использованы для ознакомления с методикой определения рыбных запасов, следует признать статьи Г. Н. Монастырского о запасах воблы (1940), о запасах сельди (1951), и о динамике численности промысловых рыб (1952), где имеется большой материал методического значения и где можно почерпнуть сведения по истории этого вопроса, т. е. ознакомиться с разными методами определения рыбных запасов. Этими статьями я и воспользуюсь, поскольку пока нет возможности предлагать каких-либо общепринятых способов и инструкций по данной проблеме.
Из русских исследователей первым серьезно подошел к теории рыбных запасов Ф. И. Баранов (1925), предложивший теорию рыбного промысла. Сущность этой теории заключается в следующем:
1. Рыбные богатства водоема определяются его кормностью.
2. Кормность (количество продуцируемого водоемом корма) и все прочие факторы остаются неизменными.
3. На рыбные запасы оказывает влияние интенсивность вылова.
4. Средняя величина довоенного улова равна годовому приросту запаса.
По Баранову, прирост рыбы в водоеме есть величина переменная и определяется тем избытком корма, который остается в водоеме за вычетом корма, потребленного живущей в нем рыбой на поддержание ее существования. Чем больше этот избыток корма, т. е. чем меньше остается в водоеме рыбы, тем больше получается прирост. Под остатком Баранов понимает разность между величиной запаса и годовым уловом рыбы.
А. В. Морозов (1932 а) считает, что о состоянии рыбных запасов можно судить, сопоставляя интенсивность промысла с количеством выловленных рыб, и придерживается такой точки зрения:
1. Запасы рыбы регулируются интенсивностью промысла.
2. Изменения запасов вызываются также и ограничительными мерами в отношении маломерной рыбы.
3. Возрастной состав рыб изменяется под действием промысла.
Г. Н. Монастырский не считает возможным пользоваться при оценке рыбных запасов ни методом Ф. И. Баранова, ни методом А. В. Морозова, как основанными на промысловой статистике без биостатистического анализа уловов.
Биостатистический метод, по мнению Г. Н. Монастырского, – «основное средство для оценки состояния запасов и для составления промысловых прогнозов».
Биостатистическим методом в числе первых пользовался А. Н. Державин (1922), по мнению которого для выводов о запасах рыб необходимо иметь: статистику уловов за ряд лет, соответствующую жизненному циклу рыб, знание возрастного состава уловов и допущение постоянства этого состава на протяжении изучаемого периода.
Г. Н. Монастырский считал, что возрастной состав используемого промыслом стада может не иметь сходства с возрастным составом всей массы рыбы, и это не обеспечивает получения правильного представления о запасах рыбы.
Тем не менее Ф. И. Баранов, А. В. Морозов и А. Н. Державин содействовали развитию учения о рыбных запасах. А. Н. Державин положил основу для биостатистического метода изучения запасов и в числе первых выступил с теорией определения запасов рыб. Ф. И. Баранов (1918) и А. В. Морозов (1932) пользовались преимущественно статистическим методом, однако все последующие ихтиологи, занимающиеся методом определения запасов, в своих расчетах в той или иной мере используют указания Державина, Баранова и Морозова.
Астраханская ихтиологическая лаборатория и другие при оценке состояния рыбных запасов пользовались: 1) статистическими сведениями об уловах, 2) данными о средней длине и весе рыбы, 3) сведениями о возрастном составе уловов (Монастырский, 1940). Перечисленные материалы необходимы для общих суждений о запасах.
Г. Н. Монастырский (1940), разобрав многие методы определения запасов рыб и прогнозирования уловов (Баранова, Морозова, Державина, Астраханской ихтиологической лаборатории, Терещенко, Фаррана, Ходжсона, Роллефсена), сделал следующие выводы:
1. Главными факторами, обусловливающими изменение запасов рыбы, являются величина поколений, рост, пополнение и убыль.
2. Пополнение запаса за счет одного какого-нибудь поколения происходит ежегодно на несколько лет. Абсолютный и относительный темпы пополнения из года в год различны и обусловлены величиной поколения и темпом роста.
3. Изменение возрастного состава стада рыбы и роста ее не зависит от интенсивности лова.
4. Биостатистический анализ уловов, понимаемый в широком смысле, имеет очень большое значение для определения запасов рыб.
Такими положениями Г. Н. Монастырский пользовался, когда излагал свои выводы о запасах и прогнозах воблы Северного Каспия (1940).
В распоряжении исследователя имелись данные об уловах воблы за многие годы. Эти данные показывали, что уловы достигают максимальной величины дважды в год: осенью и весной, что обусловлено биологическими особенностями рыбы:
осенью – при подходах к местам зимовки, весной – во время распадения льда в море и при нерестовых миграциях. Эти скопления приурочены к температуре воды менее 14-15°С.
На протяжении осени и последующей весны вобла привязана к одному и тому же району. Осенняя, достигшая половой зрелости, вобла по своему возрастному составу и длине тела не отличается от весенней, идущей на нерест. На основании этих положений, полученных за многие годы, можно по осеннему стаду воблы прогнозировать перспективы весеннего лова и предсказывать возрастной состав, средние размеры возрастных групп и навески. Далее автор обсуждает вопрос о средних пробах, которые являются основными материалами для заключения о запасах.
Необходимо брать пробы одновременно в нескольких местах, и чтобы проб было «обилие» из районов скопления. Наибольшее значение придается пробам, добываемым исследовательскими орудиями, которые могут брать рыб всех возрастных групп, встречающихся на одном месте. Такие пробы «должны всецело основываться на распределении и миграциях рыб», т. е. собирать пробы следует в то время, когда рыба распространяется на меньшей площади и образует скопления густые». Г. Н. Монастырский признает достаточной пробу в 100 экз. рыб. Осенью он рекомендует брать пробы в море один месяц- с середины октября до середины ноября; в это время образуются наиболее значительные скопления воблы. Периоды распадения льда и миграции воблы длятся 1,5 месяца, с конца марта до первой половины мая. В это время надо также брать пробы. Установлено, что достаточно брать пробы один раз в пятидневку. Кроме того, пробы собирались на нескольких судах, чтобы сбором материалов охватить весь район скоплений воблы. На судах пробы брались ежедневно, но обрабатывали по 1-2 пробам за пятидневку.
В то же время брали пробы (по 100 проб в пятидневку) в течение всей путины, на тех пунктах, куда поступали уловы с основных мест лова.
Каждая проба подвергалась такой обработке: измеряли длину, брали чешую для определения возраста рыб, определяли пол и зрелость половых продуктов, а также вес рыбы. По пробам устанавливали возрастные группы выловленной рыбы. Так как «основную часть уловов воблы составляет зрелая вобла, необходимо условиться называть всю совокупность зрелых особей промысловым стадом или промысловым запасом».
«Промысловое стадо воблы состоит: 1) из остатка стада после нереста предшествующей весной; 2) из пополнения особями, впервые достигшими половой зрелости, и 3) из остатка стада, состоящего из особей, пропустивших один-два сезона нереста».
Исходным пунктом в вопросе о запасах рыбы (в данном случае воблы) автор считает возрастной состав промыслового стада. Процентные отношения зрелых особей в пробах запасов являются показателями пополнения промыслового стада». Большое значение в методике определения промысловых запасов имеет анализ нерестовых колец на чешуе рыбы. Рыба с одним или несколькими нерестовыми кольцами составляет так называемый остаток, а не имеющая нерестового кольца зрелая рыба-пополнение промыслового стада (запаса).
Наряду с пополнением идет и убыль стада. Убыль должна бы равняться пополнению, но качественно убыль не соответствует пополнению.
Зная зависимость промыслового стада от величины пополнения, роста и убыли, можно составить характеристику этого стада и указать перспективы лова на ближайший год.
Из приведенных выше соображений Г. Н. Монастырского ясно, что для того, чтобы судить о запасах и давать прогнозы уловов рыбы, необходимо иметь такие материалы:
1. Статистические данные об уловах за несколько лет.
2. Массовые данные о средней длине и весе рыбы.
3. Массовый возрастной материал со средними длинами возрастных групп.
4. Наблюдения за нерестовыми площадями и условиями жизни молоди рыб.
Полезные результаты при этом может дать мечение рыб.
Несмотря на огромные и многолетние материалы, которыми пользовался Г. Н. Монастырский, и несмотря на тщательный их анализ, он все же приходит к выводу, что в настоящее время, когда нет почти никаких количественных данных об естественной смертности воблы, «мы можем лишь обсуждать вопрос о порядке величины запасов воблы». Можно говорить об увеличении или об уменьшении запасов на ближайшее время (на следующую путину).
В 1951 г. опубликована другая статья Г. Н. Монастырского, где исследователь сделал такие заключения по методике учета и прогноза численности рыб:
1. Устанавливается состав нерестовой популяции (зрелой рыбы) на основании средних проб, собранных неотбирающими, отцеживающими орудиями лова.
2. В результате учета рыб с нерестовыми марками определяется соотношение между пополнением (впервые созревающие рыбы, без нерестовых марок) и остатком (зрелые особи, с нерестовыми марками).
3. Определяется возрастной состав пополнения.
4. Определяется темп роста основных возрастных групп нерестовой популяции.
5. Оценивается мощность поколений на основе учета мальков (сеголетков) исследовательскими орудиями лова.
6. Определяется плотность популяции на местах зимовки и в период зимовальных миграций как исследовательскими, так и промысловыми орудиями лова.
7. Дается оценка относительной численности нерестовой популяции.
8. Устанавливается коэффициент корреляции между фактическими уловами, плотностью популяции и численностью поколений, а затем путем экстраполяции определяется величина возможного улова.
Повторяем, что, несмотря на серьезнейшую разработку вопросов методики определения состояния и прогнозирования запасов рыб, выполненную Г. Н. Монастырским и другими исследователями Советского Союза и зарубежных стран, пока нет надежного метода такого определения. В вышедшей в 1955 г. книге П. Г. Борисова и А. С. Богданова «Сырьевая база рыбной промышленности СССР» имеется глава о методике оценки состояния запасов рыб и о прогнозах ожидаемых уловов. В этой главе основательно, хотя и кратко, разобраны практиковавшиеся попытки многих авторов найти способы определения запасов рыб (работы Т. С. Расса, И. И. Месяцева, А. А. Шорыгина, В. Г. Майского, А. Н. Смирнова, Ф. И. Баранова, Н. Л. Чугунова, А. Н. Державина, Н. А. Маслова, Г. Н. Монастырского, Т. Ф. Дементьевой, Н. П. Танасийчука и др. и ряда иностранных ихтиологов).
Простой перечень авторов показывает, что интересующийся данным вопросом научный работник должен овладеть познаниями обширной литературы прежде, чем приступить к решению задачи о запасах стада или отдельных видов рыб того или иного водоема. Надо иметь в виду и то, что много полезного можно найти и в тех работах, которые не преследовали особой цели определения запасов, но посвящены изучению биологии и промысла рыб. П. Г. Борисов и А. С. Богданов справедливо указывают, что для оценки состояния запасов рыб необходимо исследовать большое количество факторов, влияющих на запасы. Авторы перечисляют следующие главнейшие из таких факторов: урожай молоди рыб, скорость роста, возрастной состав нерестовой популяции, величины пополнения и убыли. Относительно прогнозов возможных уловов те же авторы говорят, что прогнозы можно давать лишь по отношению к тем видам рыб, над которыми ведутся многолетние ежегодные наблюдения, позволяющие проследить колебания численности исследуемых рыб, которые «облавливаются промыслом со значительной интенсивностью». «Для рыб, запасы которых еще слабо используются промыслом, прогнозы лишены смысла, и для них большее значение имеет общая ориентировочная оценка состояния запасов для определения возможности развития или организации их промысла».
Все исследователи, занимающиеся проблемой определения запасов рыб и прогнозирования уловов, считают, что одним из основных факторов, определяющим величину рыбных запасов и оптимальных уловов, является естественная смертность рыб. Однако вопрос этот до самого последнего времени оставался недостаточно изученным.
Большое значение в связи с этим имеют исследования П. В. Тюрина (1962, 1963), предложившего оригинальную методику определения коэффициента естественной смертности, разработанную им главным образом в результате разностороннего анализа возрастной структуры стад ряда промысловых рыб. Здесь можно лишь кратко изложить содержание его весьма важных в методическом отношении исследований.
Под коэффициентом естественной смертности автор понимает отношение части популяции, отмершей в течение года от разных причин (не считая промысла), к величине всей популяции в начале года, выраженное в процентах. Равным образом коэффициент вылова – отношение части популяции, выловленной в течение года, к величине всей популяции в начале года, выраженное тоже в процентах. Коэффициент общей смертности – сумма коэффициентов естественной смертности и вылова.
Развивая и уточняя положения Ф. И. Баранова о закономерности убыли стада промысловых рыб от вылова и естественной смертности и С. А. Северцова о закономерностях динамики численности животных, учитывая также, что в средних возрастах естественная смертность ниже, чем в младших и старших, П. В. Тюрин пришел к выводу о возможности определения коэффициента естественной смертности у рыб в средних возрастах по их вероятному (теоретическому) предельному возрасту. Знание же коэффициента естественной смертности рыбы позволяет определить наименьший промысловый размер, фактический и оптимальный коэффициенты вылова, допустимый предел прилова молоди, а также рассчитать величину возможного улова популяции в данном водоеме на ближайший год.
В указанных работах П. В. Тюриным приводятся таблица w график для определения коэффициента естественной смертности рыб, и излагается методика пользования ими. Зная фактический предельный возраст рыбы, относимый к той или иной по числу экземпляров пробе, можно по таблице или графику установить вероятный (теоретический) предельный возраст рыбы, а по нему определить и коэффициент естественной смертности, свойственный средним возрастам, что особенно важно в практическом отношении, так как эти возрасты определяют основные промысловые запасы рыбы.
По мнению П. В. Тюрина, весьма важный вопрос о наименьшем промысловом размере рыбы можно решить правильно лишь с учетом коэффициента естественной смертности. От этого коэффициента зависит темп увеличения ихтиомассы популяции нарастающей, как правило, не безгранично, а лишь до определенного предела (возраста), после которого весовой прирост популяции с возрастом становится меньше, чем убыль от естественной смертности. В связи с этим П. В. Тюрин предлагает
наименьшие промысловые размеры рыб определять по кульминации ихтиомассы популяции, найденной с учетом их коэффициентов естественной смертности. При этом у мирных рыб возрастная группа, предшествующая наступлению кульминации ихтиомассы, должна вступать в нормальную эксплуатацию наравне с другими старшими возрастными группами (у хищных рыб эксплуатацию выгоднее начинать несколько раньше).
В качестве примера приводится расчет по лещу оз. Ильмень (табл. 13).
Т а б л и ц а 13
Определение наименьшего промыслового размера леща оз. Ильмень (по П. В. Тюрину, 1963)
|
Возраст, годы |
Исходная численность по возрастам, шт. |
Гибель от естественной смертности 22%, шт. |
Длина до С, см |
Вес1 экз., г |
Вес возрастной группы, кг |
|
2 |
1000 |
220 |
9,6 |
15 |
15 |
|
3 |
780 |
172 |
14,4 |
57 |
40 |
|
4 |
608 |
134 |
18,6 |
140 |
85 |
|
5 |
474 |
104 |
22,4 |
241 |
114 |
|
6 |
370 |
81 |
27,7 |
438 |
162 |
|
7 |
289 |
64 |
32,6 |
735 |
212 |
|
8 |
225 |
49 |
35,0 |
936 |
211 |
|
9 |
176 |
39 |
37,9 |
1233 |
214 |
|
10 |
137 |
30 |
38,9 |
1338 |
183 |
|
11 |
107 |
24 |
41,6 |
1691 |
181 |
|
12 |
83 |
18 |
46,8 |
2150 |
178 |
Как видно из таблицы, кульминация ихтиомассы в популяции леща; наступает в возрасте 7 лет. Следовательно, промысел его следует начинать с возраста 6+, и тогда наименьшая промысловая Мера определяется в 30 см при весе около 500 г, что подтверждает правильность установления меры на леща Правилами рыболовства 1960 г.
Общие положения по проблеме естественной смертности и •практические выводы из установленных П. В. Тюриным закономерностей сводятся к следующему (Тюрин, ,1963, стр. 80-82).
«1. Коэффициенты естественной смертности у рыб весьма различны и связаны с длительностью жизни особей в популяциях видов. В данном отношении рыб можно подразделить на три основные группы.
Первая группа – рыбы весьма долговечные с поздним наступлением половозрелости. Живут свыше 30 лет. Коэффициент естественной смертности низкий-меньше 16%. Кульминация нарастания ихтиомассы (с учетом естественной смертности) наступает при массовой половозрелости видов (осетр), иногда позднее (стерлядь). Коэффициент вылова обычно выше коэффициента естественной смертности. Поэтому ограничения в промысле обязательны. Главные из них: лимитирование вылова; охрана нерестилищ и пропуск на них достаточного количества производителей; охрана от преждевременного вылова молодых возрастных групп путем установления наименьших промысловых размеров, соответствующих возрасту достижения массовой половозрелости видов.
Вторая группа-рыбы со средней длительностью жизни в пределах 15-25 лет. Массовая половозрелость наступает в возрасте (4)-5-6-7-(8) лет. Коэффициент естественной смертности от 20 до 35%. Кульминация нарастания ихтиомассы у одних видов совпадает с возрастом массовой половозрелости (судак, лещ) или даже превышает его (щука), у других видов наблюдается при достижении лишь начального полового созревания (сиговые). Коэффициенты вылова равны коэффициентам естественной смертности рыб (оз. Ильмень) или превышают их (Северный Каспий, Азовское море). Поэтому на промысел ценных видов этой группы должны существовать такие же ограничения, как и для рыб первой группы.
Наименьшие промысловые размеры мирных рыб данной группы следует устанавливать на 1 год раньше возраста, при котором вид достигает наивысших показателей ихтиомассы. У хищных рыб этой группы необходимо учитывать увеличение кормового коэффициента с возрастом. Поэтому их наименьшие промысловые размеры должны быть определены в возрасте, на 2-4 года предшествующем достижению кульминации ихтиомассы.
Третья группа-виды с очень кратким периодом жизни, скороспелые. Живут обычно от 3 до 10-13 лет. Половой зрелости достигают в возрасте l-3 лет. Коэффициент естественной смертности очень высок (95-40/о), поэтому кульминация нарастания ихтиомассы наблюдается на 1-м (снеток, тугун) или на 2-3-м (мелкая ряпушка, салака) годах жизни.
К этой группе, кроме перечисленных рыб, можно отнести ерша, окуня, плотву, уклею и многих других малоценных и сорных (обычно многочисленных) рыб.
Коэффициенты вылова этих видов обычно ниже, чем коэффициенты их естественной смертности. В связи с этим промысел на них может быть значительно повышен за счет той части популяции, которая неизбежно погибает в результате естественной смертности. Поэтому установление наименьших промысловых размеров и лимитов на вылов рыб данной группы лишено биологического обоснования. В отношении ценных видов рыб этой группы целесообразно устанавливать сроки или участки лова, ограничивающие или запрещающие вылов рыбы на нерестилищах, чтобы не подорвать уровень естественного воспроизводства, а также устанавливать наименьшие размеры ячеи в орудиях лова, отсеивающих, если это необходимо, первую возрастную группу рыб.
Промысел малоценных и сорных видов никаким ограничениям не подлежит. Однако при наличии в водоеме ценных рыб, молодь которых в значительных количествах может вылавливаться одновременно с малоценными и сорными рыбами, борьба с последними должна осуществляться преимущественно при помощи ценных хищников (судака, лососевых, щуки и др.), для чего необходимо соответствующими мерами резко увеличить их стада.
2. На основании рассмотренных примеров и расчетов по некоторым другим рыбам можно предположить, что каждой отдельной популяции того или иного вида свойственен свой коэффициент естественной смертности и свой тип динамики нарастания ихтиомассы. При этом у видов с большим ареалом южные популяции характеризуются более высокими, а северные популяции – более низкими коэффициентами естественной смертности. В связи с этим должны быть различными и наименьшие промысловые размеры рыб.
3. В зависимости от колебаний численности популяции по годам в целом и по отдельным поколениям, а также обеспеченности их кормом, темп роста у рыб может изменяться. В связи с этим могут меняться динамика нарастания ихтиомассы, возраст и размеры рыб при наступлении половой зрелости. Все это может обусловить изменение наименьших промысловых .размеров в ту или другую сторону.
4. Наименьшие промысловые размеры, определенные изложенным методом, являются минимальными, так как применение меньшего коэффициента естественной смертности неизбежно вызовет их повышение.
5. Все изложенное не может быть распространено на виды рыб, жизненный цикл которых прерывается катастрофически (дальневосточные лососи, угорь и др.)».
Исследуя далее вопрос о величине вылова промысловой рыбы при разной интенсивности рыболовства, П. В. Тюрин приходит к выводу, что оптимальный коэффициент вылова не должен превышать коэффициента естественной смертности. Однако этот вопрос более детально был исследован в следующей работе на основании анализа количественных изменений воспроизводительной способности популяции леща озера Ильмень при разных коэффициентах вылова. Эти исследования позволили установить следующее:
1. Приспособительные способности облавливаемой популяции рыбы к воздействию факторов внешней среды, в том числе и к изъятию промыслом, (имеются в виду в сравнении с необлавливаемой популяцией улучшение темпа роста, ускорение полового созревания, увеличение плодовитости) крайне ограничены и не превышают в количественном выражении коэффициента естественной смертности. Отсюда объективным критерием интенсивности вылова рыбы является ее коэффициент естественной смертности.
2. Оптимальный коэффициент вылова рыбы не должен превышать ее коэффициента естественной смертности. Следовательно, коэффициент общей смертности промысловой рыбы не должен превышать удвоенного коэффициента естественной смертности (здесь имеется в виду некоторое снижение коэффициента естественной смертности от коэффициента вылова).
3. Более высокие коэффициенты общей смертности, превышающие удвоенный коэффициент естественной смертности, ведут к снижению воспроизводительной способности промыслового вида и, как следствие, к подрыву численности стада и падению улова.
4. Контроль за интенсивностью рыболовства и состоянием запасов промысловой рыбы вполне надежно осуществляется по возрастной структуре нерестовой популяции и особенно по .наличию предельных возрастов в исследуемых средних пробах.
Считая установленные действующими правилами рыболовства нормы прилова рыб непромысловой меры заниженными и биологически необоснованными, П. В. Тюрин предлагает при определении допустимого предела прилова молоди также исходить из приспособительных свойств облавливаемой рыбы к факторам внешней среды, в том числе и к изъятию промыслом. Если Приспособительные свойства вида к изъятию промыслом равны коэффициенту естественной смертности в каждом возрасте, то вполне очевидно, что в условиях рационального промысла на оптимальном уровне, когда осуществляется весь комплекс биологически обоснованных мер регулирования, прилов молоди разных возрастов в количествах значительно меньших, чем коэффициент естественной смертности для каждой возрастной группы в отдельности, никакой опасности для данного вида не представляет.
П. В. Тюрин считает, что удобнее всего предел допустимого прилова молоди устанавливать в процентах по счету от улова данного вида, как это принято в правилах рыболовства. Тогда допустимые нормы прилова молоди могут быть определены следующим образом:
а) в том случае, когда охраняемый вид малочислен (в годовом улове составляет менее 10%) и, кроме промысла, испытывает пресс со стороны других рыб, процент прилова молоди разных возрастов по счету от улова данного вида допустим в размере его коэффициента естественной смертности;
б) если охраняемый вид многочислен (в годовом улове составляет более 20%) и господствует над другими рыбами, процент прилова молоди разных возрастов по счету от улова данного вида допустим в размере 1,5 коэффициента естественной смертности;
в) в отношении весьма малочисленных или особо ценных видов, отличающихся низкой воспроизводительной способностью (осетровые, лососевые, некоторые сиговые и др.), необходимо запрещение всякого прилова и требование немедленного выпуска в воду всех жизнеспособных особей непромысловой меры.
Сказанное можно пояснить примерами. Лещ в оз. Ильмень является многочисленным и господствующим видом. Коэффициент естественной смертности у него 22%. Допустимый предел прилова молоди по счету от улова леща в промысловых орудиях равен 22х1,5=33%, а соотношение молоди и мерной рыбы в улове равно 33 : 67.
Лещ в Ладожском озере малочислен, и там эти соотношения уже будут иными (22 : 78).
В табл. 14 приводятся примерные допустимые нормы прилова молоди ценных промысловых рыб по П. В. Тюрину.
П. В. Тюрин (1963) излагает также основные принципы оценки состояния рыбных запасов и нормирования уловов с учетом положений, разработанных главным образом Г. Н. Монастырским, Т. Ф. Дементьевой и Г. В. Никольским.
Он приводит примеры оценки запасов и определения возможного вылова некоторых рыб в северо-западных озерах путем использования предлагаемой методики: с учетом коэффициентов вылова, естественной смертности, общей смертности, численности вылавливаемых рыб по возрастным группам, ихтиомассы общей и по отдельным поколениям.
Излагая эту методику, П. В. Тюрин указывает на необходимость широкой проверки ее на практике, что даст возможность дальнейших уточнений этого сложного вопроса. В частности, требуется регулярный сбор материалов, который позволит определять численность по возрастным группам всей выловленной в водоеме рыбы и одновременно уточнять исходные коэффициенты общей и естественной смертности.
Таблица 14
Примерные пределы допустимого прилова молоди ценных промысловых рыб в условиях рационального промысла
|
Группы рыб |
Теоретический предельный возраст, годы |
Коэффициент естественной смертности . % |
Соотношение допустимого процента прилова молоди и процента мерных особей по счету от улова охраняемой рыбы |
|
|
малочисленные виды (менее 10% в годовом улове) |
многочисленные виды(более 20% в годовом улове) |
|||
|
|
100 60 35 |
Менее 5 71 |
Подлежат выпуску в воду |
|
|
Рыбы средней долговечности |
30 25 20 15 |
16 20 25 35 |
16 :84 20:80 25:75 35:65 |
24:76 30:70 37:63 52:48 |
|
|
13 10 8 5 4 3 |
40 50 60 80 90 95 |
Ограничению не подлежат |
|
Особо ценные материалы по данной проблеме могут быть получены в результате полного учета численности и ихтиомассы в обрабатываемых химическим методом озерах, а также путем математического моделирования динамики численности популяций. Первые опыты в этом направлении уже осуществлены.
Примеры определения фактической численности рыб, ихтиомассы и возможных уловов в обработанных химическим методом озерах
В СССР широкое применение получает химический метод рыбохозяйственного преобразования озер, разрабатываемый лабораторией акклиматизации ГосНИОРХ. Его сущность, практическое и теоретическое значение изложены в ряде работ автора этого метода-Ё. В. Бурмакина, в частности, в его статье «Химический метод рыбохозяйственного преобразования озер» (1963).
После обработки полихлорпиненом (ядовитое вещество, которое широко применяется е сельском хозяйстве) озера полностью освобождаются от рыб. Обрабатывают таким способом водоемы с малоценными видами. Затем озера удобряют (если необходимо – предварительно известкуют) и по истечении срока детоксикации заселяют ценными видами рыб, а иногда и кормовых организмов.
В результате такого комплекса мелиоративных мероприятий рыбопродуктивность озер в умеренной зоне СССР повышается с 10-20 кг/га (до обработки) до 100-200 кг/га.
Ценность этого метода заключается и в том, что он обеспечивает разрешение ряда весьма важных теоретических вопросов озерного рыбного хозяйства. При хорошо поставленном учете и комплексных исследованиях можно точно определить численность рыб, общую ихтиомассу (вес всей рыбы), ихтиомассу видов рыб и отдельных их поколений, предельный возраст и возрастную структуру популяций, а на основании этого – выявлять закономерности внутривидовых и межвидовых отношений, а также закономерности, определяющие размеры ихтиомассы и численности рыб, рыбопродуктивность озер различных типов, обосновывать допустимые выловы по видам и др.
Эти данные можно будет использовать и при ведении рыбного хозяйства на таких озерах, которые нельзя обработать химическим методом.
Е. В. Бурмакин и Л. А. Жаков (1961) излагают результаты определения численности, ихтиомассы и естественной рыбопродуктивности в небольшом озере Тюленьем (Ленинградская обл.), которое было обработано полихлорпиненом.
Площадь оз. Тюленьего-1,83 га, в нем обитал, только окунь. Его популяция при обработке озера учтена полностью и оказалась состоявшей из 3181 экз. общим весом 81,02 кг в пересчете на площадь-1738 экз/га или 44,3 кг/га.
Была взята проба для определения возрастного состава и роста окуня. Это позволило рассчитать его численность по возрастным группам, средний годовой прирост ихтиомассы каждой группы в отдельности и всей популяции в целом.
Суммарный средний годовой прирост ихтиомассы окуня, который авторы называют естественной рыбопродуктивностью озера, составил 26 кг для всего водоема или 14,2 кг/га.
Из этого количества на долю неполовозрелых особей, пополнения и «рекрутов», т. е. на возрастные группы 0+-3+, пришлось 9,1 кг/га и на взрослых особей (4+-8+) -5 кг/га.
Обращает внимание преобладание окуня возрастной группы 3+. Особей этого возраста было 1494 шт., или около 47% •численности всей популяции. Наименьшая численность была- у сеголетков – всего 78 шт., или 2,5%.
Такое же соотношение численности возрастных групп окуня было отмечено ранее и в других окуневых озерах – Окуневце и Плавущем (Бурмакин, 1960).
Л. А. Жаковым раньше было установлено, что в оз. Тюленьем в течение ряда лет откладывалось приблизительно одно и то же количество икры (400-450 кладок-). Поэтому низкая численность окуня младших возрастных групп, выявленная при обработке озера, не могла быть объяснена низкой его урожайностью. Авторы приходят к выводу, что это является следствием уничтожения молоди более крупным окунем: «как бы много личинок не появилось… они, по-видимому, тотчас же и почти полностью уничтожаются рыбами предыдущих поколений».
Однако по истечении времени, равного приблизительно длительности жизненного цикла окуня (в оз. Тюленьем до 8- 9 лет), «наступает такой момент, когда численность рыб «каннибалов» настолько сокращается, что они не могут существенно сократить новое поколение и это новое поколение занимает в течение нескольких последующих лет ведущее положение в популяции».
Е. В. Бурмакин и Л. А. Жаков приходят к выводу, что в окуневых озерах сеголетки окуня хотя и могут быть многочисленными, но таких случаев значительно меньше, чем преобладания окуней старших возрастных групп. Таким образом, были выявлены особенности структуры и динамики численности стада окуня и рыбопродукционные возможности окуневых озер.
Заметим, что заключение о 8-9-летней цикличности динамики численности популяции окуня в оз. Тюленьем было впоследствии подтверждено В. В. Меншуткиным (1964), применившим метод математического моделирования популяции рыб на примере окуня этого озера.
Е. В. Бурмакин (1960) приводит данные и о результатах химической обработки и учета рыбы в озере Жемчужном (тоже в Ленинградской области). Площадь этого озера 68,81 га. Оно было населено 9 видами рыб: окунем, ершом, плотвой, щукой, лещом, налимом, красноперкой, карасем и пелядью (выпущенной в озеро сеголетками). Общая численность рыб составила 469,7 тыс. экз., ихтиомасса 3913,8 кг (6826 экз/га и 56,9 кг/га). По численности и ихтиомассе преобладали окунь, ерш и плотва. В пересчете на 1 га численность этих рыб округленно составила 6800 экз., ихтиомасса – 52 кг.
На основании выполненных работ Е. В. Бурмакин высказал мнение, что «ихтиомасса однотипных озер при сходных ландшафтных условиях и одинаковом составе рыб является довольно постоянной величиной». Для лесных окуневых озер Северо-запада СССР ихтиомасса составляет 42-45 кг/га, а ежегодный ее прирост или допустимый вылов- 14-15 кг/га в год. Мелководные мезотрофные с признаками дистрофии озера, как Жемчужное, населенные разными рыбами с преобладанием окуня, ерша и плотвы, имеют, вероятно, ихтиомассу, равную 55- 56 кг/га, возможный годовой вылов 20-25 кг/га, т. е. тоже незначительный. К тому же при существующих (естественных) межвидовых отношениях в уловах всегда будут преобладать малоценные рыбы. Для повышения в таких озерах уловов и улучшения их качественного состава необходимы специальные комплексные мероприятия по реконструкции ихтиофауны.
Г. П. Руденко (1962) в результате исследования популяций окуня в 5 озерах (включая оз. Тюленье) пришел к выводу, что ихтиомасса окуневых ацидотрофных озер является величиной более или менее постоянной, но «при условии одинаковой стадии развития озер и одинаковой фазы развития популяции, а не только при сходстве ландшафтных условий». Продуктивность озер меняется по годам и зависит от фазы развития популяции и величины ихтиомассы, в случае преобладания молоди – рыбопродуктивность возрастет.
Математическое моделирование динамики численности рыб
Биологические основы принципа математического моделирования в ихтиологии разработаны Г. В. Никольским (1963), который указывает на перспективность и совершенную необходимость применения такого метода при исследовании динамики численности популяций рыб.
Большой интерес и весьма существенное методическое значение в этом отношении представляет работа В. В. Меншуткина (1964) «Исследование динамики численности рыб на основе представления популяции в виде кибернетической системы».
Автор указывает, что методы математического моделирования популяций можно разделить на две группы. Одна группа представляет модели с непрерывным временем и с использованием для вычислений дифференциальных уравнений (Баранов, 1918 и др.). Но трудности при решении сложных дифференциальных уравнений не позволяют отразить с нужной полнотой сложную приспособительную сущность взаимодействий популяции со средой. Вторая группа – модели с дискретным временем, когда популяции рассматриваются через промежутки времени, обычно через один год. Такой метод моделирования допускает построение моделей неограниченной сложности, что как раз и характерно для биологических систем. Элементы подобного моделирования имеются в работах Г. В. Никольского (1961, 1963), Ю. Е. Лапина (1961), а также А. Н. Державина (1922) и Н. Л. Чугунова (1935), посвященных проблеме определения запасов рыб.
В. В. Меншуткин исследовал динамику популяции методом кибернетического эксперимента. Это первый опыт применения кибернетики в ихтиологии. Появление быстродействующих электронных вычислительных машин обеспечивает возможность выполнения большого объема вычислительных работ, а быстрое развитие кибернетики-теоретическую базу для моделирования.
Моделью послужила упоминавшаяся уже популяция окуня из озера Тюленьего, исследованная Е. В. Бурмакиным и Л. А. Жаковым. Вычислительные работы производились на машине БЭСМ-2М Сибирского энергетического института Сибирского отделения АН СССР, а подробное описание алгоритма системы (программы) было дано В. В. Меншуткиным и Л. А. Жаковым на Х Конференции по водоемам Прибалтики.
При моделировании популяции окуня из озера Тюленьего ее состояние определялось фактически установленной численностью девяти возрастных групп (0+-8+). Учитывалось изменение численности в результате «каннибализма», гибели от старости, промысловой гибели, ограниченности кормовой базы. Принималось во внимание изменение плодовитости с возрастом, а также влияние внешних факторов на гибель икры. Интервал между двумя состояниями системы был принят в один год. Дано подробное описание блок-схемы программы вычислений.
Исследования были выполнены в двух вариантах: 1) при условии отсутствия всякого промысла и 2) под воздействием на популяцию рыболовства различной интенсивности.
В первом случае независимо от первоначального состояния стада при неизменяемости условий среды (в частности, кормности водоема) и внутренних связей в стаде популяция выходит на один и тот же цикл.
В. В. Меншуткин следующим образом характеризует циклические изменения необлавливаемой популяции окуня оз. Тюленьего:
1. В состоянии минимальной численности популяция в основном состоит из неполовозрелых особей, причем количество сеголетков относительно невелико (на диаграмме в работе автора – фаза I).
2. В последующие годы (фазы II и III) увеличивается численность младших возрастов вследствие увеличения количества производителей. Одновременно намечается «провал» в численности возрастных групп 2+ и 3+, который объясняется «каннибализмом» со стороны более старших особей.
3. Этот провал наиболее характерен для состояний максимальной численности популяций, где он приходится уже на возрастные группы 4+ и 5+ (фазы IV и V).
4. Достижение малочисленными возрастными группами половой зрелости обусловливает уменьшение количества сеголетков (фазы VI и VII).
5. После гибели от старости большого числа особей наступает состояние, близкое к фазе I.
Периодичность таких колебаний структуры и численности популяций окуня составляет 6-8 лет.
При исследовании влияния на численность популяции промысла, за первоначальное, исходное ее состояние бралось такое, в каком популяция оказывается в результате ее естественного развития в течение продолжительного времени (автор брал 100 лет, считая, правда, что это излишне большой срок).
Было принято девять градаций интенсивности рыболовства- от 0,1 до 0,9′(За интенсивность рыболовства – F принято «отношение числа выловленных особей к наличной численности облавливаемой части популяции при условии применения неселективных орудий лова»), при семи разных размерах ячеи сетей, соответствующих отбору окуня определенных возрастных групп. Средние годовые уловы подсчитывались при всех возможных комбинациях интенсивности рыболовства и размера ячеи (на это потребовалось немногим больше 1,5 ч работы машины).
Получены весьма интересные результаты. Например, при наибольшей интенсивности рыболовства (0,9) и использовании сетей, отбирающих трехлетков (2+), популяции окуня через 5 лет отлавливается полностью; при интенсивности 0,7 и тех же сетях – через 25 лет. Максимальные выловы отмечены при сетях, отбирающих пятилетков (4+) и интенсивности рыболовства 0,9. При лове сетями, рассчитанными на отлов окуня старых возрастов (например, 6+), общая численность популяции увеличивается по сравнению с ее численностью в необлавливаемом водоеме. Это объясняется ослаблением каннибализма вследствие вылова крупных особей. Наоборот, лов мелкоячейными сетями, отлавливающими окуня младших возрастов, влечет за собой снижение общей численности популяции.
Работа В. В. Меншуткина иллюстрируется графиками и диаграммами, характеризующими полученные им результаты по исследованию динамики численности окуня в необлавливаемом
озере и под влиянием воздействия промысла различной интенсивности.
Автор отмечает и недостатки своего эксперимента: состояние кормовой базы в озере условно принималось за постоянную величину (в виду отсутствия соответствующих данных), рост рыб – зависящим только от возраста, т. е. без учета других факторов и т. п. Вместе с тем он указывает, что при предлагаемом методе моделирования популяций в виде кибернетических систем вполне возможны дальнейшие усложнения программы моделирования многими данными по биологии и экологии рыб, которых в его распоряжении не было.
Метод В. В. Меншуткина, несомненно, имеет большое перспективное значение, поскольку он дает возможность анализа основных биоэкологических факторов, обусловливающих динамику численности рыб. Это весьма убедительно доказывается в кратко рассмотренной здесь работе В. В. Меншуткина.
Глава XI
ПРИНЦИПЫ СОСТАВЛЕНИЯ РЫБОПРОМЫСЛОВЫХ КАРТ
Определение термина я содержания научно-промысловой карты
Промысловая карта-одна из наиболее выразительных форм обобщения и передачи в практику нужных для рыбного хозяйства сведений о промысловых объектах и промысловых водоемах.
Промысловую карту мы определяем как графическое отображение главнейших сведений, относящихся к самому промысловому водоему и к промысловым организмам, с учетом перспектив промыслового хозяйства. Такое определение указывает на характер и количество тех элементов, какие должны быть выражены на карте. Все элементы промысловой карты служат цели – содействовать рациональному использованию промысловых богатств данного водоема в соответствии с его естественными условиями.
Употребляя здесь термин «промысловые богатства», мы имеем в виду не только рыб, но и другие промысловые объекты: беспозвоночных, млекопитающих и даже водные растения. Существующий многие годы термин «рыбный промысел» при настоящем состоянии промысла, когда значительную долю его составляют, помимо рыб, многочисленные животные организмы и даже растения, своевременно или изменить, или расчленить. Можно употреблять вместо общего термина «рыбный промысел» термин «водный промысел» или «промысел водных продуктов», а выражение «рыбный промысел» применять только к добыче рыбы. На основании приведенной оговорки в понятие «промысловой карты» включается весь водный промысел (рыбный, промысел беспозвоночных и млекопитающих). Но мы подробно будет говорить о рыбном промысле, потому что последний имеет наибольшее хозяйственное значение, да и в ихтиологической области накоплено большое количество нужных для нашей темы примеров и фактов.
Сущность содержания промысловой карты можно представить и виде четырех кругов, из которых три (внутренних) вписаны в четвертый.
Центральный (первый) круг означает сведения о промысловых объектах (или промысловом объекте) – это круг промысловой биологии. Второй (средний) круг включает сведения по гидрологии и гидробиологии водоема – круг промысловой гидрологии и гидробиологии. Третий круг – круг сведений о современном состоянии промыслового хозяйства – преимущественно сведений, относящихся к добыче водных промысловых организмов. Четвертый (наружный) круг включает данные о перспективах хозяйства.
Второй круг находится в наиболее сложных взаимоотношениях с другими кругами. Он дает те сведения о водоеме, какие нужны промыслу для овладения промысловыми ресурсами, и те сведения об условиях жизни промысловых объектов, какие необходимы промыслу для обеспечения наиболее рационального построения всего промыслового хозяйства, включая и заботы о воспроизводстве промысловых богатств.
Упрощенная, чрезмерно сжатая передача гидрологических и гидробиологических сведений или полное их игнорирование здесь недопустимы. Недопустимо также, чтобы научно-промысловая организация, составляющая промысловую карту, занималась самым детальным и отвлеченным от промысла гидрологическим и гидробиологическим изучением водоема. Такое изучение должно идти по иному плану и для других целей.
Содержание научно-промысловой карты можно выразить более коротко: научно-промысловая карта – картографический очерк промыслового хозяйства водоема. Нельзя такой картографический научно-промысловый очерк исполнять по одной форме для всех водоемов. Для одних водоемов можно ограничиться одной картой (если малый водоем изображается в большом масштабе, или когда степень изученности водоема слаба), для других водоемов нужно и можно дать целый промысловый атлас. При этом нельзя забывать, что наша промысловая карта во всем своем содержании динамична: биологические и промысловые элементы, постоянно изменяющиеся, нельзя представить в каких-то застывших, неизменных формах. В картах должна быть отражена сезонность. Составленную карту нужно дополнять в последующие годы.
Разработке вопроса о составлении рыбопромысловых карт по внутренним водоемам в начале 30-х годов стал уделять значительное внимание Всесоюзный научно-исследовательский институт озерного и речного рыбного хозяйства. В 1934 г. была опубликована специальная статья И. Ф. Правдина «Указания к составлению научно-промысловых карт». Сотрудниками ВНИОРХ были разработаны рыбопромысловые карты по Ладожскому озеру (Правдин к повторно Шуколюков и Титенков), Онежскому (Покровский, Александров), Балхашу (Петров и Виноградов), Ильменю (Тюрин). Псковско-Чудскому водоему (Тюрин), по 10 крупным озерам Карелии (Александров, Беляева, Покровский, Смирнов и др.), по Финскому заливу (Михин и затем повторно Телегин), по Енисею от г. Красноярска до р. Кувейки (Подлесный), по Цимлянскому водохранилищу (Лапицкий) и по ряду других водоемов. Все эти карты переданы для использования рыбохозяйственным организациям, которые оценивают их весьма положительно.
В настоящем разделе Руководства автор не ставит задачи – дать подробный разбор существующих карт, которых к настоящему моменту опубликовано значительное количество. Мы рассматриваем те карты, где имеются методические указания.
В 1955 и 1956 гг. Зоологическим институтом АН СССР и Тихоокеанским институтом рыбного хозяйства и океанографии выпущены хорошо оформленные и со многими научными и промысловыми показателями «Атласы океанографических основ рыбопоисковой карты Южного Сахалина и южных Курильских островов». Многочисленные карты (в двух томах Атласа), исполненные в крупных масштабах, изображают распределение планктона и бентоса по отдельным участкам исследованных районов, грунты, течения, температуры, солености, подводные ландшафты, сезонное распределение животных и многое другое. Основы построения этих карт близки к нашему пониманию научно-промысловых карт, хотя дальневосточные карты имеют более целенаправленное наименование как карты рыбопоисковые. Хотя еще нет общепризнанных схем построения подобных карт, но нужда в таких схемах признается, что особенно наглядно проявилось в Совещании по вопросам поведения и разведки рыб в 1953 г. В статье Л. С. Бердичевского, помещенной в Трудах названного совещания, дан критический анализ очень многих промысловых карт, и автор справедливо делает такое замечание: «Самое понятие «промысловые карты» до сих пор четко не определено. Каждый автор вкладывает в это понятие свое содержание». Сам Бердичевский под промысловыми картами понимает «такое графическое изображение результатов научно-исследовательских и поисковых работ, которое дает совершенно четкое представление о характере распределения, местах и сроках наибольших скоплений в водоеме промысловых рыб или других объектов промысла, показанное в динамике».
Картирование общегеографических сведений о водоеме
Промысловые водоемы различны по своим физическим и биологическим условиям (моря, их участки, озера и реки). Мы имеем в виду составление промысловой карты преимущественно для озер, но стремимся, чтобы наши указания были достаточно общи и приложимы при работах по составлению промысловых карт разнообразных водоемов.
Первое, что необходимо иметь составителю промысловой карты-обычную географическую карту в масштабе 1, 2, 3, 5 или 10 км в 1 см. С этой карты должны быть сняты на кальку или на хорошую восковую бумагу копии, которыми исследователь должен пользоваться как макетами при полевых работах на водоеме (в момент сбора материалов о водоеме) и при работах в лаборатории (при картировании сводных данных о водоеме). Эти же макеты используются и при сборе других материалов промысловой карты.
Лучше, если рабочие основы карт будут изготовлены типографским способом.
Отдельные, наиболее существенные в промысловом отношении участки водоема следует вычерчивать особо, чтобы более детально отображать на карте все нужные сведения. Для такого деталирования удобно применять пикетажные книжки (с миллиметровой бумагой), которыми широко пользуются топографы.
На географическую основу карты наносят сетку квадратов или кварталов, которая поможет вести промысловую ориентировку. Размеры квадратов избирают условно, причем следует рекомендовать придерживаться общей градусной сетки, т. е. линий меридианов и параллелей. Для этого градусы меридианов и градусы параллелей разделить на равные части, но при этом следует иметь в виду, чтобы сетка квадратов давала квадраты, площадь которых по крупным водоемам с размерами свыше 100 км2 не превышала 25 км2. По меньшим водоемам площади квадратов должны быть меньших размеров.
Для удобства разбивки на квадраты лучше брать так называемые меркаторские карты, где меридианы и параллели даны прямыми линиями; такими картами пользуются судоводители. Называя нашу сетку сеткой квадратов, мы нарушаем геометрическую точность. Выставляя требование совмещения линий географических градусов или минут с линиями наших квадратов» мы точных геометрических квадратов не получим, а получим прямоугольники, и наша сетка в сущности будет не сеткой квадратов, а сеткой прямоугольников; такую сетку следовало бы назвать сеткой кварталов.
При игнорировании линий географических градусов (что допустимо) промысловые кварталы можно нанести любого размера, но необходимо, чтобы раз нанесенные кварталы (их размер и нумерация) всегда и для всех были обязательны. Поэтому сетку квадратов или кварталов необходимо вычертить еще до начала полевых работ. Исследователь, составляющий карту кварталов, должен самым тщательным образом обдумать и обсудить с участием хозяйственников (с обязательным участием представителей судовождения) как масштаб карты промыслового водоема, так и масштаб кварталов.
Такие карты в настоящее время имеются.
Промысловая характеристика водоемов обычно не ограничивается характеристикой только данного водоема: чаще встречается необходимость давать характеристику хотя бы некоторых участков бассейна, наиболее связанных с основным водоемом. Промысловая карта Северного Каспия должна иметь и дельту Волги; промысловая карта зал, Куйлюс Аральского моря должна быть с картой нижних участков Сыр-Дарьи; Амурский лиман – с низовьями Амура; Белое море – с речными промысловыми участками. Карты озер в большинстве должны также иметь изображение низовьев впадающих в них или верховьев вытекающих из них рек, потому что биология промысловых- рыб озер теснейшим образом связана с речными водами, и промысел использует как основной водоем (или его части), так и прилегающие к нему участки рек. Промысловая карта может быть составлена отдельно для реки и для озера, но редко на всем протяжении реки имеются существенные промысловые участки.
Для многих озер, особенно небольших, а также для рек существенное значение имеет общий береговой ландшафт: горы, овраги, низины, береговые валы, оползни, стоки дождевых и почвенных род, ключи, береговая растительность. Подобные сведения на карте отметить следует, но только в береговой, примыкающей к водоему зоне. Здесь, как и всюду в промысловой карте, должны употребляться условные знаки обозначений (рис.87).
Береговые горы и овраги имеют значение при стоке атмосферных осадков (дождей и тающих снегов).
Окружение водоема сухими или болотистыми низинами до известной степени определяет характер той же прибрежной зоны; прибрежные пески, прибрежные топи и т. д.
Береговые поля, имеющие склон к водоему, дают последнему удобрение. На карте нужно указать ручьи и углубления, по которым дождевая или снеговая вода поступает в водоем. Болотные ручьи, наоборот, часто приносят в водоем воды плохие, кислые, малокормные. Береговые ключи нередко имеют значение восстановителей кислорода в тех озерах, где (зимой) рыбы ощущают недостаток в этом газе. Вливающиеся в водоем реки вносят много органических веществ, которые повышают кормовое для рыб значение предустьевых участков. Именно в этих участках обычно наблюдаются скопления рыб и на кормежке и на нерестилищах. Поэтому картирование устьевых и предустьевых участков должно быть детализировано.
Береговые валы, располагаясь параллельно береговой линии водоема, свидетельствуют о намывающем действии прибоя или об усыхании водоема. Нередко такое же распределение имеют берега водоема подвержены размывам, а скалистые участки берегов определяют сравнительное постоянство береговой линии.
![]() Береговая растительность (камыши, луга, кустарники, лес) также отмечаются на карте.
Береговая растительность (камыши, луга, кустарники, лес) также отмечаются на карте.
Ознакомление с берегами водоема представляет еще и тот интерес, что признаки берегов могут служить и признаками самого водоема. В работе П. Ф. Домрачева (1923) очень убедительно изображена связь рельефа берегов с озерными глубинами. Выводы П. Ф. Домрачева о зависимости рельефа берегов от глубин хорошо изложены Д. А. Ласточкиным (1925) в его научно-популярной книжке «Стоячие водоемы (озера и пруды)». «Большое количество озер, имеющих высокие берега, относится к наиболее глубоким озерам, наоборот, низкие берега больше свойственны мелким озерам; каменистые, песчаные и глинистые берега более свойственны глубоким озерам, болотистые-мелким озерам, поэтому зарастание и заболачивание берегов в большей степени присуще мелководным озерам».
Рис. 87. Условные обозначения берегового ландшафта (по Правдину, 1939):
1 – горы; 2 – скалистые берега (обрывы); 3-оползни; 4-береговой вал (отдельный холм); 5 – овраги; 6–низина сухая «пляж»; 7-низина болотистая; 8 – камыши; рдесты; 9 – луг; 10-поля (огороды); 11-пески; 12- кустарник; 13 – лес лиственный, 14 – лес хвойный; 15-ключи; 16-ручьи полевые; 17 – ручьи болотные; 18 – каналы (канавы).
Береговая карта будет использована не только при характеристике водоема, но и при характеристике промысловой биологии и промысла. Отметка затопляемой (при высоких водах, особенно весенних) части берега позволит составить представление о местах выхода весеннемечущих рыб на нерестилища (например, щуки). С характером берега обычно связан и характер лова: на отлогих песчаных берегах удобно производить притонения невода.
Рис. 88. Карта сведений о берегах и глубинах малого озера (по Правдину, 1939);
1 – лес хвойный; 2 – лес лиственный; 3 – кустарник; 4-болото; 5-луг; 6-поле.
Пример картографирования сведений о берегах водоема представлен на рис. 88. Озеро окружено сложным рельефом: есть болота, холмы, луга, поля, леса и кустарники; болотные воды стекают в озеро. Глубины озерные подходят близко к берегам.
Картирование гидрофизических элементов
Из гидрологических элементов водоема на промысловой карте следует отметить главнейшие, имеющие связь с промыслом, а именно: глубины, грунты, температуры, течения. Эти показатели на промысловой карте должны занять одно из самых главных мест.
Чтобы представить на промысловой карте глубины водоема, надо провести большие гидрологические работы. Для промысла существенно знать расположение глубин, особенно тех мест, где развит или может быть развит промысел. Более детально глубины нужно представить в прибрежной полосе, где обычно нерестуют многие рыбы. В прибрежных местах исследователь должен дать рыбаку указания на мели, обозначая их особо, например пунктиром.
Если мы попытаемся изобразить сложную систему фарватеров Амурского лимана, которые имеют огромное значение и в биологии главных промысловых рыб (кеты и горбуши) и в судовождении, то линии изобат внесут путаницу и в без того сложную систему линий, указывающих фарватеры. На подобной карте глубины следует указать цифрами (с обозначением метров). Эти цифры по многим значительным водоемам можно взять из карт, изданных Главным гидрографическим управлением.
При составлении батиметрической карты нельзя ограничиваться промерами случайно полученными (при случайной поездке на водоем) или немногочисленными.
При составлении батиметрической карты нетрудно дать (или на отдельном листе или на полях общей промысловой карты) один или несколько профилей дна. Знания дна помогут рыбаку при его работах по установке орудий лова и при поисках рыбы.
Донные бороздины (углубления) и склоны служат местами скоплений рыб. От этих углублений зависят и придонные течения.
Не менее существенно знать грунты водоема, причем изучение грунтов для промысловой карты следует вести по двум разделам: а) физические (вернее механические) свойства грунтов и б) свойства биологические. Химический состав грунтов представить на промысловой карте, по-видимому, слишком трудно, а самые главные химические свойства грунта, которые так или иначе влияют на химический режим воды, мы можем отметить на карте при указаниях основных химических свойств воды. Биологические свойства дна водоема будут рассмотрены нами при указаниях по картированию гидробиологических элементов.
Физические свойства грунтов исследуются, возможно, подробнее, чтобы можно было охарактеризовать с этой стороны весь водоем, особенно дно промысловых участков. На карте нужно отметить условными знаками признаки грунтов: ил, песок, глина, камни подводные и надводные, галька, железная руда, ракушник.
Нашим перечнем, несомненно, не охвачено все разнообразие грунтов, которые встречаются в практике исследователя водоемов, мы перечислили только основные виды грунтов. Общеизвестно, что ил создает среду, благоприятную для развития донных организмов, которые являются источником питания рыб; пески, рудоносный слой и камни, наоборот, бедны животными. Важно выделить места, где имеются ил и железная руда, так как эти грунты поглощают большое количество кислорода.
Зная грунты, рыбаки смогут правильно использовать орудия лова или создать соответствующие приспособления, чтобы орудие не врезывалось в илистое дно или не застревало на камнях, задевая за них.
Температурный режим водоема представляет ряд сложнейших явлений, которые, с одной стороны, зависят от многочисленных и разнообразных причин (общее состояние атмосферы, характер берегов, впадающие в водоем реки, ключи), с другой стороны, сами в сильной степени влияют на жизнь водоема (на его растительный и животный мир.) Надо получить средние температуры водоема за несколько лет и за каждый месяц в отдельности. На некоторых водоемах такая работа ведется гидрометеорологическими учреждениями; ихтиологу остается только взять необходимые сведения и свести их так, чтобы они были связаны с биологией водоема. На промысловой карте, где даны сведения о промысловых рыбах, местах их скоплений (нерестовых и нагульных), местах миграций (где указываются сроки нереста рыбы, сроки ее подходов, сроки лова), нужно дать и температурные показатели.
Практически не всегда удается дать на промысловой карте изотермы по каждому месяцу, да, возможно, эти изотермы не для каждого водоема будут нужны. Так, указав на карте место и срок нерестового скопления того или иного вида рыб, можно указать среднюю поверхностную температуру, относящуюся к этому месту и к этому сроку.
Поверхностная температура определяет миграции и нерест многих рыб, но зимние скопления рыб, а также миграции и нерест рыб глубоководных требуют указаний и придонной температуры.
Хозяйственники и рыбаки учитывают указания на температуры, которыми сопровождается то или иное биологическое явление, соответствующим образом планируют свою промысловую работу. Так, рыбаки волжской дельты срок хода сельди уже давно устанавливают термометром. Температура в водоемах изменяется закономерно, и ее изменения, а вместе с тем и ход биологических факторов, как правило, можно предвидеть.
Полезно дать на карте (или в ее легенде) средние сроки покрытия и замерзания водоема, а также указать места, где лед бывает наиболее толст и где его не бывает.
Помимо средних сроков замерзания и вскрытия всего водоема, для промысла нужно знать сроки развития ледяного покрова в отдельных участках водоема.
Основоположник русских научно-промысловых исследований П. М. Книпович (1902) с особенной четкостью установил прямую зависимость распространения тресковых рыб Баренцева моря от течений.
По поводу этой связи Н. М. Книпович говорит следующее: «Карта течений объясняет нам многое о распределении животных. Особенно интересны данные относительно распределения и миграций важнейших промысловых морских рыб в зависимости с теплым течением. Промысловая рыба, и особенно треска, встречается преимущественно в области теплого течения и его ветвей и в непосредственном соседстве с ним».
Дальнейшее развитие мурманского промысла подтверждает справедливость указаний Н. М. Книповича. Тот же исследователь своими работами в 1914 г. установил подробную связь передвижений рыб в Каспийском море. Теперь эта связь с успехом используется промыслом. Мы имеем в виду установленные Н. М. Книповичем (1921) круговые течения воды в Каспийском море и связанные с этими течениями температуры воды. Тогда же (в 1914 г.) названный автор подметил, что осенью наибольшее количество осетровых рыб было в зоне главной струи
восточно-бережного течения «где температура была всего выше».
Связь передвижений промысловых рыб с течениями Каспийского моря установлена для многих видов. Сельдь весной (март-апрель) идет густыми косяками вдоль северного побережья Азербайджана (севернее Апшеронского полуострова) с юга на север против имеющегося здесь морского течения в северную часть Каспийского моря (на икрометание). Позднее получено еще более определенное указание на связь миграций каспийских сельдей с течениями вод Каспийского моря.
Связь передвижений рыб с течением рек общеизвестна: одни рыбы пользуются наиболее быстрыми струями воды, другие предпочитают тихую воду; в известные сроки рыба идет против течения, в другие сроки – по течению.
Такое же явление можно проследить и в озерах. Так, в Ладожском озере направляющееся с юга на север вдоль восточного берега течение образует в озере своего рода реку, которая и регулирует миграции рыб. Сиги и ряпушка после нереста, очевидно, сплывают по течению этой «реки» в более глубокие участки озера, где находят мало изменяющуюся температуру, около 4°С. Там они держатся всю зиму, а летом, по мере пробуждения инстинкта размножения, многочисленные стада сигов и ряпушки направляются против течения к восточному побережью. Основываясь на этих выводах, Институт озерного и речного рыбного хозяйства в 1932 г. рекомендовал организовать траловый лов сигов около восточного берега Ладожского озера. Практика подтвердила, что именно в этом районе траловый лов наиболее успешный.
Течения несут с собой и пищу для рыб. Рыбаки давно отмечали, что в озерах и морях появляются различного оттенка полосы воды, которые указывают на обилие или малое количество рыбы.
Болгарский исследователь Дренский (1920) занимался проверкой подобных наблюдений рыбаков. Болгарские рыбаки в Черном море различают: а) воду зеленоватую или синюю- прозрачную и бедную рыбой и б) воду желтую, более богатую рыбой. Дренский исследовал желтую воду и нашел, что она обильна ракообразными Copepoda, за которыми охотится скумбрия (цитируем по И. Я. Сыроватскому, 1939).
Картирование течений возможно, если изучено направление течений. Карта течений Баренцева моря, составленная Н. М. Книповичем, очень наглядна. Поток отепленной атлантической воды, выйдя из Атлантического океана в Баренцево море (между Шпицбергеном и северной оконечностью Скандинавского полуострова), образует широкую струю тепловодного Нордкапского течения, которая разливается по Баренцеву морю и, смешиваясь здесь с массами холодной воды полярных морей, расчленяется на несколько ветвей. Одна из этих ветвей так называемое Мурманское течение приближается к Мурманскому берегу и идет параллельно ему. Карту и описание течений (по Книповичу) Баренцева моря можно найти в книге Г. И. Танфильева (1931).
В перемещениях рыб большую роль играют и ветры. Ветер вызывает то или иное течение воды, а последнее вызывает передвижение рыбы. При ветрах верхние слои воды направляются по ветру («вода идет из ветра»), а нижележащие слои идут против ветра («вода идет в ветер»).
Помимо этих горизонтальных перемещений воды имеются еще перемещения вертикальные, или конвекционные, которые вызываются главным образом разностями температур слоев поверхностных и более глубоких.
Рыбы не могут относиться безучастно к ветровым и конвекционным течениям, но так как те и другие течения имеют значительную случайность, то промысловая карта, очевидно, не сможет представить подобные материалы, хотя картографирование преобладающих (например, по месяцам или по четырем основным временам года) величин этих явлений вполне доступно.
На карте промыслового водоема можно (и следует) давать знаки не только направления, но и быстроты течений, если есть данные по средним величинам течений; да и отдельные наблюдения над быстротой потока воды, приуроченные, например, к моменту массового хода рыбы или к моменту откладывания икры, представляют хороший материал для экологической характеристики связи биологии рыб с условиями среды. Пример картографирования подобных гидрологических материалов можно найти в работе К. А. Гомоюнова – Гидрологический режим бухты Патрокл в связи с метеорологическими условиями (1928.) На карте стрелками указаны направления течений, а быстрота течений обозначена цифрами, поставленными над (и под) стрелками.
Картирование гидрохимических элементов
Химический состав воды имеет большое влияние как на качественный, так и на количественный состав населяющих водоем организмов, в том числе и на рыб. Поэтому в рыбохозяйственных исследованиях большое внимание уделяется вопросам изучения химизма воды. Основными элементами химического анализа промысловых вод избираются: кислород, сероводород, окисляемость, углекислота, концентрация водородных ионов (рН), азот, фосфор, железо. В некоторых случаях (при расширенных гидрохимических работах) производится полный химический анализ воды (и полный химический анализ грунтов). Все перечисленные элементы можно нанести на промысловую карту, но при этом необходимо помнить, что карту нельзя перегружать обозначениями, так как тогда пользование ею станет затруднительно. Поэтому на промысловую карту надо наносить лишь те гидрохимические элементы, которые отражают ближайшую связь с показателями биологическими, в первую очередь с показателями биологии рыб.
Гидрохимические показатели надо давать в средних величинах (по месяцам, сезонам лова, или по временам года).
Для картирования сведений по газовому режиму водоема нужно указывать распределение кислорода, окисляемости, углекислоты.
Н. А. Мосевич (1931) провел гидрохимические исследования в Волховской губе Ладожского озера. Часть карт, составленных И. А. Мосевичем, передается (в сокращенном виде) на рис. 89. Рисунок так нагляден, что не требует никаких дополнительных пояснений: подпись под рисунком вполне исчерпывает весь смысл этой карты. Особенно интересно изображена на карте степень окисляемости: наибольшая окисляемость (больше 15 мг О2/л воды) обнаружена как раз против – коллектора, по которому приходят в Ладожское озеро сточные воды Сясьского бумажного комбината.
На промысловых картах особо следует отмечать участки с отрицательными отклонениями от нормы. Например, необходимо наносить на карту места, где содержание кислорода сильно понижено или отсутствует, что укажет рыбакам на отсутствие рыбы или на заморы. Такие заморные места нередко встречаются во многих озерах, преимущественно зимой. Параллельно с картированием газового режима надо наносить на карты грунты. Так, отметке илистого грунта, должны сопутствовать отметки содержания кислорода и степени окисляемости. Нужно отмечать также участки, где нормальный состав воды нарушен (в ущерб биологии и промыслу) деятельностью человека (очаги загрязнения).
Г. Ю. Верещагин (1927) в достаточной степени осветил роль так называемой гидрологической съемки озер. Составителям научно-промысловых карт следует ознакомиться с методом Г. Ю. Верещагина и его соавторов. Величины гидрохимических элементов они выражали арифметической разностью, получаемой при сопоставлении нормальной (100%) величины, свойственной тому или иному элементу, с величиной найденной. Например, содержание кислорода в одном случае равно 83, в другом-112%; на карте в первом случае следует показать -17% (минус 17, т. е. содержание кислорода меньше нормального на 17%), во втором случае +12% (плюс 12 – содержание кислорода превышает норму на 12%). «При графическом изображении кислородных данных мы пользовались величинами, выражающими уклонения от насыщения в процентах как наиболее характерными свободными от местных влияний температур (Верещагин и др., 1927). Эти уклонения (анормальности) имеют свои минимумы и максимумы, которые нужно знать для гидрохимической характеристики водоема. Таким же порядком отмечают и другие гидрохимические элементы.
Рис. 89. Распределение кислорода и окисляемости в Волховской губе (из Правдина, 1939, по Мосевичу):
1 – кислород; 2 – окисляемость; 3 – кромка льда. Цифрами обозначено содержание О2 в °/о к полному насыщению и окисляемость в мг О2/л.
Гидрохимические показатели на карте надо связывать с наиболее существенными явлениями биологии водоема: на местах нереста той или иной рыбы показать количество кислорода, дать рН и т. п.; то же сделать на местах миграций, заморов и т. п. Такие сведения будут дополнять и характеристику грунтов водоема: на сильно заиленных или заросших грунтах зимой бывает пониженное, а в некоторых случаях и полное расходование кислорода. Картирование гидрохимических показателей (но менее наглядно, чем на приведенной карте Мосевича) проведено Е. М. Крохиным (1937) в отношении распределения бикарбонатной С02 и кремнекислоты в реках и ключах бассейна реки Большой.
Для промысловых целей карта должна включать и сведения о солевом режиме вод, о таких веществах, как кальций, идущий на образование костей рыб, раковин моллюсков, раковин и панцирей (оболочек) раков и т. д., железо, хлор, азот и фосфор.
В некоторых озерах наблюдается чрезвычайная бедность воды солями кальция и вследствие этого бедность моллюсками, которые служат пищей таких ценных промысловых рыб, как сиги. Зная содержание кальция в озере, при рыбоводных мероприятиях будут учитывать, какой рыбой заселять водоем, чтобы обеспечить ее кормом, или примут меры по минеральному удобрению озер.
Картирование гидробиологических элементов
Гидробиологические данные, подобно данным гидрохимическим, на промысловой карте должны занять также почетное место, потому что органическая жизнь водоема определяет всю биологию водоема, в том числе и биологию рыб.
Гидробиологические данные, связанные с данными гидрологическими (физическими и химическими), и составляют то, что мы называем средой обитания промысловых объектов.
Гидробиологические данные промысловой карты должны сводиться главным образом к показателям кормности водоема.
Хозяйственнику нужно знать, что вопрос о количестве рыбных запасов, имеющихся в водоеме, о промысловой продуктивности водоема может быть разрешен только при непременном выяснении количественного и качественного состава тех организмов, которые находятся в прямом отношении с рыбами. Полезно привести поэтому поводу слова К. М. Дерюгина (1931): «В сфере рыбного хозяйства гидробиологические исследования играют первенствующую роль, выясняя общие условия жизни промысловых рыб и устанавливая продуктивность водоемов, на которой базируются важные рационализаторские рыбохозяйственные мероприятия. Питание рыб происходит за счет организмов, населяющих водную толщу (планктон) и дно (бентос) водоемов. Качественный и количественный состав этого населения определяет рыбохозяйственную ценность того или иного водоема. Не всегда изобилие количества особей является показателем питательности водоема, ибо качественный состав населения может быть иногда мало пригоден для питания рыб. Кроме того, в составе населения могут быть различные хищники и паразиты, которые играют роль вредителей в рыбном хозяйстве».
Необходимо, чтобы гидробиологи, изучая планктон и бентос, давали цепь взаимоотношений планктонных и бентосных организмов между ними самими и между рыбами.
Из гидробиологических данных на промысловой карте нужно дать указания по планктону и бентосу.
Фитопланктон (растительный планктон) и зоопланктон (животный планктон) имеют в своем развитии различные фазы. Многим водоемам свойственно летнее цветение, которое чаще всего обусловливается чрезмерным развитием водорослей (разных видов). Такое цветение при отмирании водорослей может вызвать летние заморы рыбы, рыба погибает или уходит от тех мест, где наиболее сильно выражено планктонное цветение. На карте следует указать места и сроки цветения планктона, а также места, где планктон не развит. Отсутствие фитопланктона тоже служит отрицательным показателем для рыб (и для рыбного хозяйства), так как молодь многих рыб кормится фитопланктоном.
Еще более важно для промысловой карты получить указания по зоопланктону, так как многие рыбы и во взрослом состоянии кормятся мелкими планктонными животными. Поэтому, указывая на карте места и сроки нахождения зоопланктонных скоплений, мы тем самым определяем места и сроки нахождения рыб, потребляющих этот корм.
Животные организмы, обитающие на дне водоемов (бентос) входят в пищевой рацион рыб и других промысловых животных как самый основной (и по отношению к некоторым рыбам единственный) элемент.
Личинки насекомых, рачки, моллюски – все это представители животных, которыми питаются рыбы.
Гидробиологи, подобно гидрологам, различают в водоемах три зоны: литораль (прибрежная область), сублитораль (переходная зона) и профундаль или глубинная зона. Каждой из этих зон присущ особый вид придонных организмов. При достаточной исследованности водоема нужно разграничить на карте эти три зоны и охарактеризовать их по основным преобладающим формам бентоса.
Решающее значение в вопросе о пастбищах многих пресноводных рыб имеют количества обитающих в водоемах личинок комаров, но тут требуются серьезные наблюдения над этими личинками (и в разных местах и в разные сроки). Можно вывести совершенно ложное заключение о бедности дна водоема, взяв пробу в тот сезон, когда личинки хирономид находятся в минимуме в связи с только что происшедшим вылетом.
Гидробиологические работы нужно вести одновременно с работами гидрологическими и ихтиологическими. Для гидробиолога нужно иметь, карты глубин и берегов, данные по термике и т. п., что облегчит установку мест (станций) гидробиологических сборов. Как правило, гидробиологи рекомендуют основные сборы проводить у берега и меньше – в глубинной части водоема.
С. Г. Лепневой (1950) приведена схема биологического исследования донной области озерного участка. На карте даны отметки береговых скал, камней, песков, указаны травянистые прибрежные заросли и обозначены места гидрологических и биологических станций.
Следует картировать и количественные показатели бентоса» т. е. нужно приводить числовые показатели так называемой биомассы (в г/м2 или в кг/га дна) по отношению ко всему водоему и к отдельным его участкам, так как плотность расселения донных групп животных в одном и том же водоеме бывает чрезвычайно различна.
Участки водной растительности, служащие колыбелью и приютом многим животным (здесь обычно держатся и стада молоди рыб, пасутся и мечут икру некоторые виды рыб), следует обязательно наносить на карту.
Карт с гидробиологическим содержанием у нас еще мало, но составление их крайне необходимо. Следует указать на хорошую карту И. Г. Закса (1929), в которой можно найти полезные руководящие указания применительно к задачам наших промысловых карт. М. В. Желтенкова (1939) в статье о питании воблы дает карту кормовых площадей этой рыбы. В 1938 г. в Трудах Полярного института рыбного хозяйства опубликована серия карт по составу планктона Баренцева моря (карты составлены Б. П. Мантейфелем). На картах указаны места скоплений основных планктонных организмов, главным образом тех, которые служат пищей для сельди.
3. А. Филатова в Трудах того же Полярного института сделала очень интересную попытку картирования показателей биомассы бентоса в юго-западной части Баренцева моря: на карте даются показатели биомассы бентоса в г/м2.
В 1931 г. В. Н. Никитин (1939) привел данные по среднегодовой биомассе планктона Батумской бухты Черного моря и количественные соотношения основных групп планктона представил в виде отдельных диаграмм-спектров. Такой метод следует использовать при нанесении на промысловые карты данных по кормовой характеристике промысловых водоемов.
По количеству бентоса пытаются определить и количество рыбы, используя известный коэффициент Альма (см. ниже).
Картирование ихтиологических данных
В настоящее время на промысловых картах дают цифры, которые относятся к промысловой продукции водоема. Что касается общей ихтиомассы, то первые данные об общем количестве рыбы удалось получить лишь в некоторых озерах северо-западной части СССР благодаря методу химической обработки водоемов, применяемому Е. В. Бурмакиным.
Все еще нет четкости в толковании терминов ихтиомасса, промысловая рыбная продукция, рыбопродуктивность водоема, сырьевая база и ряд ихтиологов и гидробиологов употребляют эти термины в различных смыслах. Б. Г. Иоганзен (1937) предлагает следующее понимание подобных терминов. «Ихтиомасса – все рыбное население водоема (промысловые и непромысловые рыбы всех возрастов). Сырьевой запас-общее количество рыб промысловых пород всех размеров (всех возрастов). Сырьевая база – максимальная часть запаса, допустимая к вылову, без истощения его». Термин биологическая продуктивность Б. Г. Иоганзен рекомендует заменить словами – биологическая производительность. Предлагаемые Иоганзеном термины хорошо обоснованы, но в нашей практике мы пользуемся несколько иными, которые мною сформулированы (1954) так: «Под рыбной продуктивностью водоема надо понимать численность (запасы) общего рыбного стада (промысловых и непромысловых рыб) в нем; под промысловой продуктивностью – запасы промысловых рыб; под промысловой продукцией – количество вылавливаемой или могущей быть выловленной рыбы при правильном, рациональном ведении хозяйства».
В ихтиологической литературе часто упоминается о коэффициенте Альма, выражаемом формулой F/B, обозначающей соотношение между средней рыбностью (F) и средней продукцией бентоса водоема (В). Для водоемов, где рыбное хозяйство поставлено рационально, это соотношение выражается от 1/2 до 1/4, т. е. количество (весовое) рыбы в таких водоемах равно от половины до четверти веса имеющегося в водоеме бентоса; в диких (без рационального хозяйства) водоемах коэффициент Альма колеблется от 1/9 до 1/31. Все же нужно знать, что ихтиологу при определении рыбности водоема пользоваться коэффициентом Альма приходится с большой осторожностью. Подобное предостережение удачно выразил гидробиолог А. А. Черновский (1930). «Для того чтобы пользоваться коэффициентом F/B, необходимо проделать большую работу по определению его в водоемах различных типов. В настоящее время такая работа ведется и у нас и за границей. И только тогда, когда она будет окончена, можно будет знать тип озера: зная его В-продукцию бентоса, по уже известному коэффициенту F/B для данного типа озера, определить его возможную рыбность».
Если сейчас мы не можем дать хозяйственнику карт с точным количественным определением каждого вида рыб, то все же мы можем составить карты распространения отдельных видов рыб. Составление таких карт для малых водоемов большого практического смысла не имеет, но при составлении промысловой карты бассейна, района, области, края или карты большого водоема указания ареалов (мест) или границ распространения того или иного вида рыб важно и для рыбака, и для рыбовода.
В многочисленных работах Л. С. Берга (и на русском и на иностранных языках) даны карты распространения многих рыб (1932, 1949).
Имеются полезные карты распространения рыб и в Атласе Ленинградской области и Карельской АССР (1935).
В водоеме рыбы размещаются по-разному: даже в небольших водоемах можно указать места, особо излюбленные одним каким-либо видом рыб. Эти места и должны быть отмечены на промысловой карте.
Выше довольно подробно говорилось о зависимости размещения рыб от гидрологических факторов. Если треска в Баренцевом море придерживается струй теплого течения, то на карте необходимо отобразить это – указать отепленные атлантические потоки и на них нанести значки тех рыб, которые приурочены к этим потокам:
Места и направление миграций рыб, пастбищ, нереста, скоплений взрослых и молодых рыб, места, которые рыбой избегаются – все это должно быть отмечено на промысловой карте.
Миграции рыб наиболее резко выявляются в период нереста, когда -рыбы идут на нерестилища. Рыбацкое выражение «ход рыбы» обычно подразумевает нерестовую миграцию рыб, когда рыба идет «рунно», «косячно», т. е. в массовых количествах. Морские рыбы совершают значительные миграции и не связанные с нерестом (осенний ход беломорских сельдей, ход трески в Баренцевом море, осенний ход леща, воблы и других рыб в приморских частях Волги).
Салака в Финском заливе совершает миграции в строгой зависимости от нахождения, распределения и концентрации планктона, которым она питается. Черноморская скумбрия в своих миграциях находится также под влиянием размещения планктонных организмов. В этих случаях мы говорим о кормовых миграциях.
Наконец, есть термин «скатывание рыбы», что соответствует миграции отнерестившейся рыбы.
Все названные виды миграций должны быть отражены на промысловых картах. При отметках нерестовых миграций нужно знаками указать направление путей идущей на нерест рыбы. Эти пути можно отмечать на морской, озерной и речной картах и нужно указывать все, даже незначительные речки и ручьи, куда идет нерестующая рыба.
Карт с указанием нерестовых миграций рыб много. Одна из них, на мой взгляд, очень поучительная составлена М. И. Тихим (1927) для пояснения хода ладожских лососей. По карте можно проследить, что ладожский лосось, войдя в Свирь, устремляется не во все притоки этой реки, а избирает лишь некоторые из них, что представляет большой научный и хозяйственный интерес. Гидрологи должны найти причины, почему лосось в одну реку идет, а другой рекой пренебрегает.
Рис. 90. Пути хода сельди в бухтах Японского моря в 1927 г. (по Амброзу, 1931):
1-январь; 2-февраль; 3-март; 4-апрель.
Отнерестившаяся рыба должна скатываться тем же путем, каким шла на нерест, поэтому на карте можно отмечать только сроки ската.
В Трудах Полярного института рыбного хозяйства (1938) приводится оригинальная карта распределения отдельных форм (установленных на основании строения головы) сельдей в Баренцевом море. Карта распределения сельди в некоторых бухтах Японского моря в различные месяца дана А. И. Амброзом
(1931) (рис. 90). К числу таких же полезных карт можно отнести карты, составленные В. Н. Тихоновым (1939) по зимнему распределению сельди в южной части Баренцева моря. Автор дает пять карт распределения сельдей в придонных горизонтах моря в ноябре, декабре (по материалам 1931-1937 гг.), в январе, феврале и марте (1931-1938 гг.). На картах различной штриховкой обозначены районы разной концентрации сельди:
наиболее густая штриховка указывает на наиболее густые концентрации сельди.
Подобные карты миграций и распределения воблы приводятся в статьях Г. А. Караваева (1939) и Т. Дементьевой (1939). Много карт можно найти в книге П. Ю. Шмидта «Миграции рыб» (1947).
Тем же порядком, как нерестовые миграции, следует наносить на карту и другие миграции рыб: кормовые, осенний ход на места зимних залеганий. Кратковременные передвижения рыб, едва ли целесообразно приводить на карте, хотя и тут можно дать интереснейший материал. Так, целесообразно дать розу (карту) ветровых передвижений хотя бы главнейших промысловых рыб (ряпушки, некоторых пород сигов, как лудоги во время летней жары, вероятно, кефали, сельди и др.).
Помимо миграций рыб, можно показать на карте миграции и других водных животных: закономерные миграции имеют и беспозвоночные и млекопитающие. Так, по миграции крабов у Н. П. Навозова-Лаврова (1927) имеются данные, которые могут быть картированы. По распространению дальневосточного краба имеется хорошая карта в работе японского ученого Марукава (Marukawa, 1933).
По миграциям морских млекопитающих, например по миграциям гренландского тюленя, также можно дать хорошую карту.
Карту (схему) распределения водорослей в районе полуострова Басаргина приводит Е. С. Зинова (1928). На небольшой карте Зинова дает места нахождения 11 форм морских зеленых водорослей. Если нанести на эту карту показатели глубин мест нахождения водорослей, то значение такой карты повысится.
Подобные карты даны И. Г. Заксом (1929) по донным сообществам Шантарского моря; на тех картах указаны (помимо мест нахождения водорослей): линия отливов, направление приливных течений, обсыхающие во время отливов места и изобаты. Часто одни и те же места служат нерестилищами разных рыб: там, где весной метал икру один вид рыб, осенью может отложить икру другой вид. Есть указания и на то, что на одном и том же участке, на одних и тех же грунтах одновременно откладывают икру различные рыбы: ладожский рипус выметывает икру чуть позднее сига лудоги, но на тех же самых местах, где отложена, икра этим сигом. В таких случаях при картографировании придется разграничивать подобные явления сроками или выбором наиболее важного промыслового вида.
В советской литературе имеется, значительное количество карт с указанием нерестовых мест различных рыб, и обычно асе авторы, дающие карты мест нереста, одновременно на тех же картах наносят и другие факты, предваряющие или сопутствующие нересту. На упомянутой карте М. И. Тихого указаны Места нереста ладожского лосося и пути миграций. То же видим И на карте, составленной Н. В. Горским (1932), изучавшим биологию семги, входящей из Белого моря в реку Выг.
Названные карты относятся к рекам, где места нереста рыбы можно отметить точно. Труднее очертить нерестовые площади N озерах и еще труднее в морях; однако и там нерестовые площади из года в год посещаются одними и теми же видами рыб, Которые либо откладывают икру на грунт или в грунт, либо рассеивают ее по подводным зарослям. Определить эти места в водоеме и нанести их на карту – дело не легкое, но нужное.
Нерестилища обычно бывают известны рыбакам раньше и лучше, чем исследователям, на долю которых остается выяснить более точно площади, занятые отложенной икрой, гидрологические условия нерестилищ и т. п.
На карте нерестилища можно обозначать замкнутыми кругл ми, квадратами, дугами и т. п. Небольшие площади нерестилищ, если карта не сильно загружена другими знаками, можно обозначать сеткой точек.
Пастбища рыб также характеризуются скоплениями рыб, причем часто разнообразными по видовому составу. Одних рыб данное место привлекает запасами планктона, другие находят здесь обильный бентос, хищные рыбы приходят на пастбища, чтобы кормиться мирными рыбами. По этому поводу следует привести поучительные строки, написанные Н. М. Книповичем (1902). «…Изучать жизнь промысловых животных, не касаясь Жизни других животных (и вообще организмов), населяющих те же воды, совершенно невозможно … Одни из животных являются пищей изучаемых нами рыб; от их количества, распределения, периодических изменений в этом распределении и т. п. зависит количество, распределение, переселение наших рыб. Другие служат пищею тех животных, которыми питаются интересующие нас рыбы».
Подобные взаимоотношения рыб, обусловливаемые их стремлением найти себе пищу, ихтиолог может наблюдать часто. Так, ладожские лососи передвигаются в озере в зависимости от передвижения рыб, служащих лососю пищей (главным образом от передвижения корюшки).
По нашим наблюдениям, летом, особенно в июне и июле, против восточного побережья Ладожского озера, на глубинах от до 20 м скопляется большое количество судаков, которые питаются корюшкой и сигами, в то же время судак скапливается и против западного берега (против селения Коккорево), где он охотится за молодью сига, которая здесь держится, находя обильный корм (ракообразных). В южной части Ладожского озера есть очень большие нагульные скопления и других рыб: ерш скопляется против селения Кобона, лещ – против селения Дубно, стада окуня легко обнаружить (в огромных количествах) в Волховской губе. Несомненно, что такие скопления можно подметить в каждом водоеме. Язь выходит на кормежку в летние луговые поймы, образуемые после выпадения сильных дождей (места этих пойм нужно отметить на промысловой карте), в реках есть сомовьи, лещовые, окуневые и другие места, где та или иная рыба держится годами, находя здесь корм и другие благоприятные условия.
Пастбища рыб на промысловой карте удобно показывать штриховкой разного размера и разного направления.
Скопления свойственны не только взрослым рыбам, но и молоди. Скопления молоди нужно знать потому, что это резервы рыбных запасов, кроме того, по скоплениям (личинок и мальков), рыб можно определить места икрометания многих рыб. Отметив на промысловой карте скопление молоди, надо оградить эти участки от облова, сделать заказниками или заповедниками.
Картирование сведений о рыбах (как и вообще о промысловых животных) закончим указанием, что в промысловых водоемах все чаще и чаще образуются такие участки, которых рыба избегает или которые грозят рыбе даже гибелью: мы имеем в виду загрязнение промысловых вод.
Отравленные и сточные воды промышленных предприятий для рыбы гибельны. Рыба, избегая отравленных вод, изменяет пути своих миграций, нерестилища и пастбища, а будучи застигнута этими водами, гибнет. Нарушаются и места лова рыбы. Поэтому при составлении промысловой карты необходимо отметить места загрязнений.
На реках и озерах есть места, сильно засоренные сплавом леса (корой, щепой, утонувшими деревьями), которые до засорения служили нерестилищами для рыбы. Такие места также следует отмечать на промысловой карте. Даже в море бывают очаги заражения промысловых вод. Они обнаружены, например, близ лесопильных заводов в Ковдинской губе Белого моря.
Мы говорили о картировании материалов, иллюстрирующих распространение рыб в пространстве, но мало знать пути передвижений рыб, нужно указывать сроки этих передвижений. Часто слышится, что хозяйственник вовремя не подготовился к встрече рыбы, опоздал или вовсе прозевал рыбу. Причиной таких упущений служит незнание сроков хода и нереста рыб.
На промысловых картах необходимо указывать сроки, когда промысел на данном месте не должен производиться. Казалось бы, удобно говорить о так называемых запретных сроках там, где будут даны указания по составлению карт по добыче рыб. Однако мы считаем более целесообразным говорить об этом именно в разделе о рыбах, т. е. в части промысловой биологии. И в самом деле, только там запрет справедлив и нужен, где он обоснован биологически.
При определении запретных сроков с точки зрения охраны рыбных запасов исследователь должен тщательно взвесить и обдумать многие смежные вопросы по биологии рыбы и ее промыслу. Для одного водоема лов, например, щуки в период ее нереста нужен не только в смысле увеличения добычи рыбного мяса, но и в смысле борьбы со щукой, как с нежелательным конкурентом для других рыб, представляющих для данного водоема большую хозяйственную ценность, чем щука. В других водоемах и щуку необходимо включить в число объектов, подлежащих охране. Тот же подход, разнообразный в зависимости от местных условий, следует применить и в отношении установления запретных сроков для лова других рыб.
Аналогичное значение имеет установление запретных мест. Известно, что даже в Каспийском море установлены рыболовным законодательством запретные для лова места.
Лучшей помощью с научно-исследовательской стороны является картирование таких участков, которые отвечали бы в наилучшей степени требованиям рационального заказника или заповедника. Рыбные заповедники могут преследовать различные цели, но все они должны служить увеличению рыбных запасов. Заповедник следует избрать на месте нереста рыбы, или на месте миграций рыб на нерест, или на месте скоплений молоди и т. д. На карте соответственно отмечается нерестовый, миграционный или мальковый заповедник или заказник. Некоторую сложность имеет вопрос о миграционных заповедниках, но и они могут принести немаловажную пользу в деле охранения рыбных запасов. Приведем такой пример: нерестовые участки рыбы лежат в верховьях реки или в озерном или морском заливе, куда промысловая рыба проходит определенными, нередко даже совсем узкими протоками и речками. Может случиться, что данная рыба требует забот об охране ее размножения (особенно часто это требуется в отношении нерестилищ лососевых). Если нерестилища лежат в верховьях речки, то никакого труда не стоит перегородить речку там, где рыба проходит на нерестилища, и выловить таким путем все стадо, стремящееся на икрометание. Столь дикий, безрассудный способ лова рыбы (именно драгоценного лосося), к сожалению, применяется до сих пор. В таких местах миграции разумно установить не только временный заказник, но и постоянный заповедник. Следовательно, эти места должны быть нанесены на карту.
Рис. 91 дает интересный опыт картирования сведений о распределении сельдей различного возраста. Работа исполнена А. И. Амброзом (1931).
Темп роста, хорошо известный для многих промысловых рыб, нанести на карту, очевидно, невозможно, но промысловая карта должна быть снабжена сводными табличками или диаграммами, иллюстрирующими темп роста главнейших (двух-трех) промысловых видов. Этого легко достигнуть, если дать точную табличку роста в показателях длины и веса данной рыбы из разных водоемов. Путем такого сравнения хозяйственник без всяких пояснений сделает правильную оценку рыбы и даже водоема.
![]() Нужная, но не достаточно полно понятая хозяйственниками область исследования болезней рыб, несомненно, заслуживает внимания при составлении промысловых карт. Важно отметить такие очаги заражения рыб, через которые заражается население. Работы проф. В. А. Догеля и его сотрудников показали, что есть водоемы, прибрежное население которых почти сплошь заражено солитерами. Источником этого заражения являются рыбы, причем одни рыбы местного водоема заражены сильно, другие слабо, а третьи могут вовсе не быть заражены. На карте следует отметить места, где рыбы наиболее заражены, или дать к карте список тех рыб, которые представляют для населения опасность глистных заболеваний.
Нужная, но не достаточно полно понятая хозяйственниками область исследования болезней рыб, несомненно, заслуживает внимания при составлении промысловых карт. Важно отметить такие очаги заражения рыб, через которые заражается население. Работы проф. В. А. Догеля и его сотрудников показали, что есть водоемы, прибрежное население которых почти сплошь заражено солитерами. Источником этого заражения являются рыбы, причем одни рыбы местного водоема заражены сильно, другие слабо, а третьи могут вовсе не быть заражены. На карте следует отметить места, где рыбы наиболее заражены, или дать к карте список тех рыб, которые представляют для населения опасность глистных заболеваний.
Рис. 91. Распределение сельдей разных возрастов в заливе Петра Великого и прилегающих районах (по Амброзу, 1931).
Материалы, относящиеся к рыбоводству, к воспроизводству рыбных запасов, также не должны пройти мимо внимания составителя промысловых карт. Своевременная отметка на карте мест выпуска рыб и указание времени выпуска помогут правильному наблюдению и учету результатов рыборазведения.
Перечисленные вопросы не исчерпывают всего разнообразия тем, но главные ихтиологические вопросы нами здесь затронуты.
Картирование сведений о современном промысле
Места лова рыбы и других водных объектов обычно совпадают с местами миграционных, нерестовых или кормовых скоплений, т. е. с теми участками, которые должны быть закартированы как элементы ихтиологические. Однако не все подобные места рыбных скоплений известны рыбакам. Исследователь должен указать рыбакам такие места лова, которые наилучшим образом отвечали бы и биологическим условиям водоема и чисто промысловым требованиям. Часто существующие места лова хороши в смысле удобств техники лова, но не хороши по биологическим условиям (чрезмерный облов нерестилищ, полное преграждение путей миграций и т. д.). Ихтиолог должен нанести на промысловую карту те места лова, какие соответствуют только что указанным требованиям.
Карту с местами лова целесообразно объединить с картами грунтов и глубин, здесь же следует указать и препятствия, мешающие лову (отдельные камни или гряды камней, засоренность, задевы и т. д.), чтобы можно было расчистить тони.
Большое значение мы придаем определению общей площади всех облавливаемых мест водоема, это имеет решающее влияние на определение промысловой рыбной продукции данного водоема. Если безоговорочно принимать рыбную продукцию данного водоема как частное, полученное от деления всего среднего годового улова рыбы на общую площадь (в га) водоема, то тем самым мы приходим к преуменьшенной продукции тех водоемов, которые имеют необлавливаемые площади. Поэтому необходимо определить общую облавливаемую площадь и сопоставить ее со средним годовым уловом, причем показатель этого соотношения, очевидно, будет нуждаться в присвоении ему нового термина, равносильного термину промысловой рыбной продукции облавливаемой площади водоема.
Места лова связывают с местом жительства рыбаков. Это особенно полезно делать, когда на карте представлен целый водоем (или большой участок) и когда указаны местные прибрежные рыбацкие поселения.
Карту с местами лова сопровождают указанием сроков лова. Карта по местам лова сигов в Волховской губе Ладожского озера представлена на рис. 92. Если составить такую карту для всего Ладожского озера, то хозяйственники легко получат представление о полном распределении и ходе лова рыбы, т. е. мы дадим в руки хозяйственникам необходимый для них календарь рыболовства. Отсюда прямой переход к картированию мест и сроков лова по отдельным промысловым объектам.
Существенным элементом содержания промысловой части карты служат материалы по орудиям лова. Они чрезвычайно разнообразны и многие из них носят узко местный ‘характер, но это разнообразие и эту локальность нужно считать неиссякаемым источником для рационализации и реконструкции техники лова. Для промысловой карты, очевидно, следует ограничиться общей обычной группировкой орудий лова, т. е. указывать следующие категории орудий: невода (разных систем), сети (разные), мережи и крючки. Для этих основных категорий нужно установить постоянные обозначения. Подобные карты уже опубликованы, например, в статье В. В. Покровского и А. Ф. Смирнова (1932).
В Трудах Научного института рыбного хозяйства (1927) С. В. Аверинцев дал очень наглядную промысловую карту Баренцева моря, где нанесены: глубины, поток теплого течения, рыболовные банки с выделением на них мест, наиболее богатых рыбой, места лова ярусом (крючком), промысловые рыбы (треска, пикша, морской окунь, морская камбала и палтус) и сезоны (по месяцам) лова рыб (Аверинцев, 1927).
Получилась карта, не перегруженная излишними подробностями и вместе с тем представляющая разнообразные элементы (гидрологические, ихтиологические и чисто промысловые) .
![]() В рыбопромысловом хозяйстве водоема количественные показатели (определение числа рыбаков и орудий лова, уловов рыбы – весового и ценностного) составляют основу экономики и занимают ответственное место в рыбопромысловых исследованиях. Но указать одни эмпирические цифровые данные, например, о ловцах и об уловах, это только одна часть большой работы. Другая часть, более важная, должна дать ответ на вопросы: какое количество улова приходится в среднем на каждого ловца, каждое хозяйство, каждое орудие лова, на каждую единицу площади водоема; какой валовой и чистый доход (в среднем) имеет каждое хозяйство и т. д. Экономисты имеют в своем распоряжении прекрасно разработанную методику для “выполнения такой работы и техническое оформление экономических выводов, имеющее за собой давний опыт, достойно изучения и подражания со стороны биологов. Каждому из нас известны превосходные диаграммы, картограммы и графики, которые с поразительной наглядностью и простотой иллюстрируют различного рода статистические и экономические выводы. В области промысловой экономики и даже обычной рыбопромысловой статистики методика графического изображения осваивается слабо.
В рыбопромысловом хозяйстве водоема количественные показатели (определение числа рыбаков и орудий лова, уловов рыбы – весового и ценностного) составляют основу экономики и занимают ответственное место в рыбопромысловых исследованиях. Но указать одни эмпирические цифровые данные, например, о ловцах и об уловах, это только одна часть большой работы. Другая часть, более важная, должна дать ответ на вопросы: какое количество улова приходится в среднем на каждого ловца, каждое хозяйство, каждое орудие лова, на каждую единицу площади водоема; какой валовой и чистый доход (в среднем) имеет каждое хозяйство и т. д. Экономисты имеют в своем распоряжении прекрасно разработанную методику для “выполнения такой работы и техническое оформление экономических выводов, имеющее за собой давний опыт, достойно изучения и подражания со стороны биологов. Каждому из нас известны превосходные диаграммы, картограммы и графики, которые с поразительной наглядностью и простотой иллюстрируют различного рода статистические и экономические выводы. В области промысловой экономики и даже обычной рыбопромысловой статистики методика графического изображения осваивается слабо.
Рис. 92. Карта мест лова сигов в Волховской губе
Ладожского озера (по Правдину, 1939):
1-неводной лов разной рыбы;
2-лов сига лудоги и рипуса;
3-лов сига парового; 4-лов волховского сига;
5- лов крючками сига, судака, сырти и др.
Статистические сведения хорошо объединить с данными биологической статистики.
При выяснении одного из главнейших вопросов научно-промысловой работы,- вопроса об определении сырьевых запасов, необходимо знать не только весовые количества вылавливаемой (или могущей быть выловленной) рыбы, но прежде всего возрастной состав ее. Поэтому сбор статистических данных об уловах необходимо построить таким образом, чтобы были включены сведения о возрастном составе уловов. Обычно используют средние пробы уловов, т. е. данные биостатического анализа, которым занимаются биологи. Но если бы в промысловых записях указывали (хотя бы приблизительно) процент вылова молоди рыб основных промысловых видов рыб, эти сведения дали бы много полезного при составлении промысловых карт.
Число рыбаков и орудий лова нужно изображать так же, как и сведения по уловам рыбы: в одних случаях – на промысловой карте, в других – на приложениях к ней.
На промысловой карте следует указать места и характер подсобных учреждений рыбного хозяйства: места приемки и обработки рыбы, а также пути транспортировки рыбы.
Сведения о перспективах хозяйства
Научному работнику приходится участвовать и даже руководить работами по реконструкции существующего или по организации нового рыбного хозяйства. Это занятие также требует картографических материалов. Выбор места для будущего рыбхоза не может быть основан без карты. Мало назвать водоем, где намечается организация хозяйства,- нужно изучить и представить на карте соседние водоемы. В озерном крае, например, придется дать целый бассейн, выявить и картировать общий ландшафт местности, указать ближайшие транспортные пути и т. п. При такой карте хозяйственник или хозяйственное учреждение, организующее рыбный совхоз или колхоз, избавится от многих недоразумений и ошибок, которые возможны при поспешных решениях.
На карте следует запроектировать и те изменения в природных условиях водоема, какие потребуются при организации хозяйства: в одних водоемах, может быть, нужно поднять уровень и увеличить водную поверхность, в других – провести мелиорацию, третьи – обогатить кормами для рыб и т. п. Одним словом, и здесь промысловая карта необходима. Составление планов хозяйства без всестороннего научно-промыслового исследования, выразителем которого является промысловая карта, неизбежно осложнит организацию хозяйства.
Сведения о перспективах промыслового хозяйства должны быть выводами из всей работы по изучению рыбного хозяйства того или иного водоема, и составление этих сведений должно основываться на научных данных. В этот раздел нужно вносить сведения о реконструкции не только хозяйства (акклиматизация, рыбоводство), но и самого водоема (вопросы мелиорации, гидростроительства и т. п.).
Все элементы промысловой карты должны служить единой цели: содействовать рациональному использованию промысловых богатств водоема в соответствии с его естественными условиями.
Промысловая карта не должна быть громоздкой, необходимо стремиться, чтобы промысловая карта водоема занимала немного места. В условиях же речного и морского рыболовства, а также для крупных озер может быть принята и форма атласа.
Пояснительный текст в виде легенд (на самих картах и в виде отдельного текста) должен быть краток, ясен и точен, словом, отвечать поставленным выше задачам промысловой карты. Выполненной при этих условиях промысловой картой исследователь подведет итоги всем научным работам, исполненным на водоеме, а сама карта явится лучшим способом внедрения в промысловое хозяйство совокупности знаний, необходимых для этого хозяйства.
В заключение к этому разделу можно высказать следующее:
1. Несмотря на большой интерес к составлению научно-промысловых карт, пока еще никем не составлена настоящая промысловая карта, которая отвечала бы всем главнейшим требованиям, предъявляемым к такой карте со стороны науки и промышленности. При составлении промысловых карт всегда нужно учитывать, что многие элементы (и в первую очередь промысловые элементы) карты не могут быть постоянными. Промысловая карта динамична, как динамичны и жизнь водоема и промысловое хозяйство.
2. Промысловая карта должна включать в себя обязательный минимум сведений о водоеме, о промысловых рыбах и промысловом хозяйстве. Нельзя подменять промысловую карту картами отдельных промысловых материалов.
3. Обширные официальные обзоры по рыболовству, сопровождающие и поясняющие карту, сильно понижают практическую ценность ее, делая пользование такой картой трудным; но нельзя рекомендовать и другую противоположную крайность, текст должен быть небольшим, даваемым или при самих картах или в особом пояснителе.
4. На карте должны быть представлены не только качественные, но и количественные показатели, например, отмечая места и степень концентрации рыб, нужно выяснить и привести данные об уловах на этих местах, определяя уловы или на одно орудие лова или на 1 ч работы орудием (на единицу рыболовного усилия).
Неудачи при составлении карт должны не ослаблять, а увеличивать наше внимание к промысловым картам. Вспомним, какой длительный эволюционный путь пройден обычными географическими картами, прежде чем они приняли тот вид, который имеют теперь. Научно-промысловые карты также должны пройти свои стадии совершенствования.
Глава XII
ПРИМЕНЕНИЕ ВАРИАЦИОННОЙ СТАТИСТИКИ В ИХТИОЛОГИЧЕСКИХ РАБОТАХ
Математический метод, или метод вариационной статистики в последние 25-30 лет широко применяется в области биологических наук, особенно в области ботанической и зоологической систематики. Этим же методом пользуются и ихтиологи, преимущественно при работах по систематике рыб, хотя с таким же успехом математические вычисления применимы для обработки и биологических материалов (возраст, плодовитость, упитанность, линейные и весовые размеры рыб, количество выловленных рыб и вообще всюду, где требуется нахождение и сравнение средних величин). Однако никогда не нужно забывать, что вариационная статистика это – один из многих методов научной работы, и пользование им не должно носить только механический характер. При установлении морфологического диагноза рыб, чему наиболее содействует математический метод, во-первых, принимается во внимание обычно не один, а несколько признаков, отличающих описываемую форму от других, во-вторых, для математических обоснований диагноза нужен большой материал исследуемых особей и, в-третьих, так называемая дифференция признаков, находимая по формуле ![]() (пояснение см. ниже), по отношению к рыбам, у которых абсолютно точное измерение тела и его частей почти невозможно, может быть признана существенной при показателе больше, чем 3 и 3,5. Для лучшей гарантии достоверности подмечаемого расхождения пластических признаков следовало бы признать показатель не 3, а 5, а для признаков мало изменяющихся (главным образом для счетных малоизменчивых признаков) можно принимать и показатель 3 или 3,5. Предлагаемый показатель 5 математическими расчетами не обоснован, и его принимаем только как гарантию получения наиболее достоверных выводов. Здесь рассматривается приложение математического метода преимущественно к обработке измерений, получаемых при работах по систематике рыб. Понятно, каждый работающий в иной, чем систематика рыб, ихтиологической специальности вполне сумеет пользоваться нижеследующими указаниями и при своих занятиях.
(пояснение см. ниже), по отношению к рыбам, у которых абсолютно точное измерение тела и его частей почти невозможно, может быть признана существенной при показателе больше, чем 3 и 3,5. Для лучшей гарантии достоверности подмечаемого расхождения пластических признаков следовало бы признать показатель не 3, а 5, а для признаков мало изменяющихся (главным образом для счетных малоизменчивых признаков) можно принимать и показатель 3 или 3,5. Предлагаемый показатель 5 математическими расчетами не обоснован, и его принимаем только как гарантию получения наиболее достоверных выводов. Здесь рассматривается приложение математического метода преимущественно к обработке измерений, получаемых при работах по систематике рыб. Понятно, каждый работающий в иной, чем систематика рыб, ихтиологической специальности вполне сумеет пользоваться нижеследующими указаниями и при своих занятиях.
Значение индексов и модальной варианты
Подробные промеры рыб своей конечной целью имеют определение наиболее характерных, выраженных в средних величинах, признаков различных групп рыб; и эта цель может быть достигнута при помощи соответствующего математического анализа полученных просчетов и промеров. У рыб слишком мало постоянных признаков, которые не зависели бы от различных физиологических и внешних условий, поэтому только промеров недостаточно. Общая конституция рыб резко отличается от конституции других позвоночных. У рыб рост костей происходит иначе, чем у других животных. Позвоночник рыб растет весьма продолжительное время, хотя в разные периоды жизни темп роста неодинаков: у молодых – быстрее, у старых – медленнее. Собственно мы не знаем, когда заканчивается рост рыбы в длину. Также точно неизвестно, до какого возраста растут и другие части тела рыб. Голова, глаз быстро растут в молодости, а потом рост этих частей становится менее заметным, отсюда и принято говорить об относительно малом глазе, малой голове старых рыб. При изучении рыб приходится иметь дело с рядом признаков, и по времени и по темпу роста изменяющихся.
Приведем пример: положим, что мы желаем знать, какую длину имеет хвостовой стебель воблы (Rutilus rutilus caspicus). Берем 100 рыб и штангенциркулем измеряем у каждой особи длину хвостового стебля. Промеры (выраженные в мм) у рыб размером от 100 до 300 мм колеблются от 20 до 40 мм, т. е. в пределах 21 числа. Постоянства в цифрах нет. Причина такого непостоянства ясна: длина хвостового стебля находится в прямой зависимости от длины тела, является функцией длины тела: растет тело рыбы в длину,- растет, хотя и в неравномерной зависимости от общего роста тела, и хвостовой стебель. У мелких рыб хвост короче, у крупных длиннее, но это «короче» и «длиннее» не дает нам право сказать, что у воблы длина хвостового стебля бывает столько-то миллиметров. Если возьмем рыб более мелких или более крупных,- чем взятые в первый раз, размеры хвостового стебля будут совершенно другие.
Более наглядные результаты можно получить, если непосредственно измеренные величины хвоста выразим в индексах, т. е. цифрах, показывающих отношения найденных цифр к основной величине тела – к его длине. Если выразим длину хвостового стебля воблы в процентах длины тела, то ряд цифр уменьшится с 21 до 7 (принимая целые проценты, что для данного примера достаточно). Мало говорившие до этого цифры теперь приобретают значительную ценность: мы утверждаем, что длина хвостового стебля воблы колеблется от 18 до 24% длины тела. Результат получился уже более определенный, но при нашем представлении о вобле, как о подвиде (subspecies) плотвы, склонном давать свои местные формы (установлены локальные формы воблы для Астрабадского залива, Куры и др.), индексы нас тоже не могут удовлетворить.
Есть еще величина, на которой систематики нередко основывали и основывают свои вычисления. Это так называемая преобладающая или модальная величина, короче, как теперь принято называть, – мода. Моде приписывают значение определителя наиболее часто встречающейся величины признака. Длина хвостового стебля измеренных нами 100 особей взрослой воблы, будучи выражена в процентах длины тела, колеблется, как было упомянуто, между 18 и 24%: у одних особей длина хвостового стебля равна 18% длины тела, у других-19, у третьих-20 и т. д. до 24%, но количество особей в каждой группе неодинаково, а именно: длина хвостового стебля у 7 особей составила 18, у 9-19, у 26-20. у 38-21, у 13-22, у 6 – 23 и у 1 особи-24%.
Расположим приведенные индексы более наглядно:
|
% длины тела |
Число особей |
|
18 |
7 |
|
19 |
9 |
|
20 |
26 |
|
21 |
38 |
|
22 |
13 |
|
23 |
6 |
|
24 |
1 |
|
Число рыб |
100 |
Модальная величина равна 21%, потому что эта величина свойственна наибольшему числу исследованных особей (именно 38 особям).
Но мода сравнительно хорошо характеризует признак лишь в том случае, когда перед исследователем проходит большой и вместе с тем довольно однородный материал, как в приведенном примере. В нашем примере мода находится как раз в середине всего ряда величин признака (длины хвостового стебля): три величины (18, 19, 20) менее и три величины (22, 23 и 24) более модальной величины (21). Таким образом, мода (21%) очень близка к медиане (20, 71), которая делит весь ряд пополам, т. е. таким образом, что налево и направо от нее находится ровно половина всего числа особей ряда (100:2=50). Однако случаи, подобные приведенному, встречаются далеко не всегда.
Пример. А. И. Рабинерсон (1925), сравнивая число жаберных тычинок у сорокских и кандалакшских сельдей, приводит такой ряд:
Число жаберных тычинок
сорокских сельдей …. 60 61 62 63 64 65 66 67 68 69 70 n
Число случаев …….. 1 3 8 24 29 27 32 24 10 3 1 162
Модой здесь следует считать 66, медианой приближенно 65, средняя арифметическая величина будет 65,06.
Точно медиану можно определить, если представить ряд в виде ряда классовой вариации, т. е. с границами классов (см. ниже), и под верхней границей каждого класса подписать число особей до нее – иначе говоря, если составить так называемый налетный ряд. В данном случае границы первого класса, где число жаберных тычинок 60, будут 59,5 и 60,5, границы второго класса 60,5-61,5 и т. д. и ряд примет, следующий вид, причем под ним мы подпишем и налетный ряд:
Число жаберных тычинок .. 59,5 60,5 61,5 62,5 63,5 64.5 65,5 66,5 67,5 68,5 69,5 70,5 п
Количество случаев………….. 1 3 8 24 29 27 32 24 10 3 1 162
Налетный ряд ………………………. 1 4 12 36 65 92 124 148 158 161 162 162
Медиана в данном ряду должна отсечь 81 случай налево от себя и 81 случай направо (162/2 =81): очевидно, она располагается в пределах класса с границами 64,5-65,5 (с серединой 65), причем верхняя граница класса 65,5 отсекает на11 случаев больше, чем это требуется для медианы (92-81=11) Так как в данном классе размещается 27 особей (случаев), то при равномерном их распределении одна особь займет 1/27 классового промежутка, а 11 особей займут 1Х11/27 = 0,41 классового промежутка, и чтобы получить медиану, от верхней границы класса необходимо отнять это число: Med![]() (округлено до одного знака после запятой). То же число мы получим, если при решении используем нижнюю границу класса 64,5, тогда Med = 64,5+1(81-65)/27 = 64,5+1×16/27 = 64,5+0,6 = 65,1.
(округлено до одного знака после запятой). То же число мы получим, если при решении используем нижнюю границу класса 64,5, тогда Med = 64,5+1(81-65)/27 = 64,5+1×16/27 = 64,5+0,6 = 65,1.
Нередко получаются и такие ряды, где расхождение моды, медианы и средней выражаются значительно резче.
Нужно знать не только моду, но и весь возможный размах колебаний данного признака: его правую, наибольшую, и левую, наименьшую, величины, а эти последние могут и не получиться путем непосредственных промеров, или, как принято говорить, эмпирическим путем. Ниже мы увидим, что приведенный размах длины хвостового стебля (от 18 до 24%) еще не есть полный размах: здесь мы не учли, что возможны и большие отклонения от середины. Такой учет необходим, иначе мы не сможем отличить по этому признаку одну форму рыбы от другой, близкой к первой. Такой учет произвести нетрудно, если пользоваться методами вариационной статистики. Нужные для биологов сведения хорошо изложены во многих руководствах.
Вариационные ряды
Прежде всего, необходимо научиться составлять так называемые вариационные ряды и производить анализ этих рядов для определения основных необходимых для работы ихтиолога величин. Мы остановимся главным образом на технической стороне вычислений, так как ознакомление с биометрикой по существу должно вестись иным порядком.
В примере с длиной хвостового стебля воблы числа: 18, 19, 20, 21, 22, 23, 24-ряд изменчивости взятого признака, и такая изменчивость, найденная количественным путем, путем промеров и просчетов, носит название изменчивости количественной; ряд цифр изменчивости от минимального до максимального значения называется вариационным рядом, а самые члены этого ряда в данном случае 18, 19, 20, 21, 22, 23, 24 принято называть вариантами, причем различают варианты классовые и варианты целые.
С классовыми вариантами чаще приходится иметь дело, когда мы для определения признака прибегаем к измерению. В этих случаях приходится совершенно искусственно разбивать полученные варианты на группы или классы, соединяя в один класс все величины от и до такого-то предела. Так как большинство признаков рыб – признаки пластические, характеризуемые преимущественно измерительными величинами и выражаемые процентными соотношениями, то ихтиологу-систематику чаще всего приходится иметь дело с вариантами классовыми.
Процентные соотношения обычно выражают с точностью до десятых процента. Разбивая варианты на классы, мы должны написать вариационный ряд иначе, чем делали до этого. Принято писать так: 17,5-18,5-19,5-20,5-21,5-22,5-23,5-24,5
В первый класс мы включили все варианты свыше 17,5, но не свыше 18,5 т. е. 17,6; 17,7; 17,8; 17,9; 18,0; 18,1; 18,2; 18,3; 18,4 и 18,5, Следующий класс начинается с 18,6 и содержит все варианты до 19,5 включительно. Таким образом, разница между границами классов, называемая интервалом класса, или классовым промежутком, равна 1%. При такой разбивке вариантов на классы все они с любым десятичным знаком подпадут в какой-либо класс нашего ряда. Необязательно писать вариационный ряд с пятью десятыми, можно с любым десятичным знаком, например 17,1-18,1-19,1-20,1-21,1-22,1- 23,1-24,1 и т. д.
В этом ряду разница между границами каждого класса так же равна 1%. В первый класс попадают варианты с 17,2 и до 18,1 включительно; второй класс начинается с 18,2 и до 19,1 включительно и т. д. Обычно принято писать ряд с 0,5 (17,5-18,5-19,5 и т. д.).
Серединой класса, или типичной его вариантой, будет варианта, равная полусумме крайних значений класса; например, Серединой первого класса нашего первого вариационного ряда будет величина, равная полусумме крайних членов этого класса
17,64+18,5 = 35,3 = 18,05,
2 2
середина первого класса второго ряда.
(17,2+18,1):2 = 35,3:2 = 17,65.
Вот какой вид примет вариационный ряд, выражающий длину хвостового стебля воблы в процентах длины тела.
Классы, см ……………17,5-18,5-19,5-20,5-21,5-22,5-23,5-24,5 n
Число особей или
частота вариант …………….. 7 9 26 38 13 6 1 100
Число особей – числа, выражающие частоты при классовых вариантах, пишутся под серединой классов. Здесь каждый класс выражен единицей (одним процентом), т. е. от 17,6 до 18,5 включительно. При длинных рядах классы берут более широкие. Вообще дробление ряда на классы произвольно, но лучше брать нечетное число классов.
Например, нужно составить вариационный ряд длины тела рыб размером от 2 до 35 см. Если бы за класс мы избрали одну единицу (1 см), то ряд получился бы очень протяженный и, возможно, с несколькими модами; удобнее взять больший интервал, или промежуток класса, например, 5 см. Тогда получим такой ряд:
Классы, см…… 0 – 5 – 10 – 15 – 20 – 25 – 30 – 35 п
Частота…………… 4 10 30 70 27 15 1 157
Середина каждого класса здесь равна также полусумме его крайних членов: например, середина I класса (1+5) :2 = З.
Приведенный пример относится к случаю измерения длины рыб целыми сантиметрами, поэтому и классы выражены в виде целых единиц, разница между границами двух соседних классов равна 5 см: в первый класс внесены рыбы длиной тела от 1 до 5 см включительно.
Если возьмем широкий ряд с показателями, аналогичными тем, какие даны в примере, относящемся к длине хвостового стебля, то вариационный ряд будет иметь выражение, представленное в таблице показателей высоты анального плавника у самки горбуши. Высота анального плавника у самки горбуши в процентах длины основания А:
Классы, % 67,5 – 72,5 – 77,5 – 82,5 – 87,5 – 92,5 – 97,5 – 102,5 n
Частота 1 11 6 7 17 7 1 50
Ширина класса равна 5 единицам (%):
67,6 68,6 69,6 70,6 71,6
67,7 68,7 69,7 70,7 71,7
67,8 68,8 69,8 70,8 71,8
67,9 68,9 69,9 70,9 71,9
68,0 69,0 70,0 71,0 72,0
68,1 69,1 70,1 71,1 72,1
68,2 69,2 70,2 71,2 72,2
68,3 69,3 70,3 71,3 72,3
68,4 69,4 70,4 71,4 72,4
68,5 69,5 70,5 71,5 72,5
Первый класс начинается с 67,6 и оканчивается 72,5. Целые-варианты, когда их немного, выражаются только целыми единицами без объединения их в группы (классы). Целыми вариантами мы характеризуем обычно меристические или счетные признаки рыб: число чешуи в боковой линии, число лучей в плавниках, число жаберных тычинок и т. п.
Вариационный ряд целых вариант, когда ряд небольшой, имеет такой вид:
Число мягких лучей в D (у воблы)
Целые варианты …… 8 9 10 n
Частота …….………. 2 88 9 99
При большом количестве вариант их объединяют в классы. Частоту при целых вариантах указывают против вариант. Составление вариационных рядов технически рекомендуем производить следующим образом.
Прежде всего находим максимальную и минимальную величины составляемого ряда и этим определяем его ширину.
Если ширина ряда большая, более 13-15 членов, группируем по две и более варианты в один класс. Далее разносим частоты (случаи наблюдений) по классам. Практически это выражается в разноске по классам счетных или измерительных указателей данного признака. Легче разносить показатели счетных признаков, труднее группировать измерительные показатели, которые обычно выражаются с точностью до десятых, т. е. с одним десятичным знаком). Эти показатели берут из ведомости индексов.
Индексы зачитывают по порядку, и каждый из них отмечают условным знаком в соответствующем классе нашего ряда. Та-inmmh знаками избирают точки, черточки, крестики, но несравненно более удобно отмечать таким способом: первые четыре индекса (частоты) данного класса отмечают четырьмя точками, [расположенными в углах воображаемого квадрата::, вторые четыре-черточками, соединяющими наши точки, получается фигура квадрата ?, а два следующие индекса отмечают диагоналями этого квадрата. Каждая полная фигура представляет десяток (что сильно облегчает просчет). В качестве примера разноски индексов приводим табл. 15.
![]()
![]()
![]() Таблица 15
Таблица 15
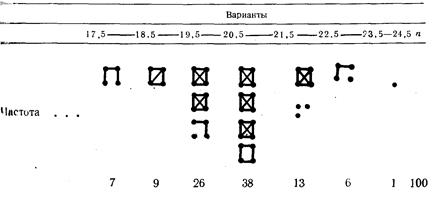
Удобен также следующий способ разнесения вариант. Варианты отмечают в так называемой разносной решетке, форма которой дана в табл. 16. Классы расположения в этой таблице по восходящим величинам, но можно располагать и по нисходящим величинам.
Ряды из целых и классовых вариант можно изображать и графически при помощи вариационных кривых, кривых изменчивости или кривых частоты. Построение простых кривых общеизвестно, к тому же ими в работах по систематике рыб пользоваться приходится мало. Заметим, что когда вариационная кривая имеет не одну, а, например, 2-3 вершины (это бывает и тех случаях, если в вариационном ряду не одна мода), то ми/кис думать, что исследуемый материал представляет собой неоднородную группу особей (быть может, сказалось влиять пола, возраста или другие причины). Графический анализ построение и внимательное рассмотрение графиков) всегда полезен как контроль математической работы. Полимодальность может быть результатом: недостаточного числа промеренных экземпляров, случайным явлением, результатом принципиальных существенных отличий. Дело исследователя в каждом конкретном случае выяснить, почему появилась одна или несколько дополнительных мод. Биологические соображения тут обычно ценнее чисто математических.
Таблица 16
|
Граница классов(от – до) |
Отметка вариант |
Частота |
|
3,1-4,5 |
/ |
1 |
|
4,6-6,0 |
/// |
3 |
|
6,1-7,5 |
//// |
4 |
|
7,6-9,0 |
////// |
6 |
|
9,1-10,5 |
//////// |
8 |
|
10,6-12,0 |
////////// |
9 |
|
12,1-13,5 |
/////// |
7 |
|
13,6-15,0 |
///// |
5 |
|
15,1-16,5 |
/// |
3 |
|
16,6-18,0 |
– |
0 |
|
18,1-19,5 |
// |
2 |
Средняя величина (М) и ее нахождение
Важно ознакомиться с основными элементами вариационных рядов и научиться эти элементы определять.
Говоря о моде, о преобладающей величине признака, мы уже успели оттенить мысль, что для характеристики признак, нужно знать его среднюю величину и все отклонения от этот средней. Средней величиной ряда является среднее арифметическое из всех его вариант. Укажем наиболее простые и на и более принятые способы этих вычислений и попытаемся дать к ним необходимые пояснения.
Формула (малоупотребительная) нахождения арифметической средней такова
![]()
где М-средняя, ?-сумма произведений, v–любая вариант, р-частота, число случаев, п-число членов ряда.
Находят сначала сумму ? произведений величины каждой варианты v на соответствующее ей число случаев р и эту сумму пят на общее число членов ряда п, на число исследованных особей.
Числовое решение приведенной формулы, если взять наш пример варьирования длины хвостового стебля воблы, следующее:
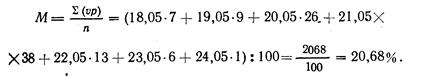
Вычисление среднего арифметического значительно упрощается, если эту величину будем высчитывать не по числам 1ВМих вариант, а по числам их уклонений от варианты, условно избранной за приближенную среднюю, т. е. будем пользоваться формулой М=А+b.
Приближенной средней, которую обозначают буквой А, обычно берут варианту с наибольшей частотой, так как она ближе других стоит к средней и чаще лежит в середине вариационного ряда.
Берем тот же пример.
Класс, см 17,5-18,5-19,5- 20,5- 21,5- 22,5- 23,5- 24,5 п
Частота 7 9 26 38 13 6 1 100
Наиболее частой вариантой является классовая варианта 1,5-21,5%, серединой этого класса нужно считать 21,05 ![]()
Поэтому приближенной средней нужно взять 21,05, т. е. А = 21,05, а варианты (собственно середины классов) влево и вправо от этой величины уклоняются на 1, 2 и 3 единицы. Левые варианты меньше, правые больше 21,05.
Варианты, меньше избранной средней величины, имеют уклонения со знаком минус (-), а варианты, больше средней, со знаком плюс (+).
Уклонения от средней М обозначаются греческой буквой а (альфа), а уклонения от А обозначаются латинской буквой а.
Тогда наш пример получит более простую (с меньшими числами) форму:
Уклонение от А(а) -3 -2 -1 0 +1 +2 +3
Частота уклонений (р) . 7 9 26 (38) 13 6 1
Высчитываем среднее уклонение, обозначая его знаком b от А по формуле
![]()
Проще будет, если запись вести таким порядком:
Уклонение (а) 1 2 3
Частота уклонений (р) -26 -9 -7
+13 +6 +1
Алгебраическая сумма
частот уклонений ( ?p) -13 -3 -6
Сумма уклонений,
помноженных на частоту
?pa -13 -6 -18 = -37
Остается -37 разделить на п, т. е. на 100, и получим сред нее уклонение:
![]()
Выражение b= –0,37% говорит о том, что настоящая средняя М от условной средней А, которая равна 21,05%, отличается на -0,37, т. е. М=А + 6=21,05%-0,37% =20,68%.
Результат получился тот же, что и в первом случае, но вы числительная работа сильно сократилась.
Если классовый промежуток не единица, как в нашем при мере, а иной, например 5, то, чтобы найти М, значение b нужно помножить на показатель этого промежутка, т. е. на 5, и при изведение сложить с А. Формула для определения М при классовом варьировании больше единицы будет: М=А+b*?, где ?-классовый промежуток, или интервал.
При малом количестве наблюдений, преимущественно при счетных признаках, и когда, кроме средней, не требуется отыскивать других вариационных элементов, не надо составлять вариационный ряд, а среднюю можно отыскивать простым (на счетах) арифметическим способом: складываем варианты и сумму делим на количество наблюдений (особей). Например, определить среднее число тычинок на первой жаберной дужке \ сунского сига (взято 15 особей):
|
Номер п/п |
Число тычинок |
Номерп/п |
Число тычинок |
Номер п/п |
Число Тычинок |
|
|
1 |
37 |
6 |
33 |
11 |
32 |
Сумма тычинок 489 |
|
2 |
32 |
7 |
37 |
12 |
32 |
Число наблюдений 15 |
|
3 |
34 |
8 |
33 |
13 |
29 |
Средняя = 489:15 = 32,6 тычинок |
|
4 |
32 |
9 |
34 |
14 |
32 |
|
|
5 |
31 |
10 |
31 |
15 |
30 |
При этом всегда необходимо указывать минимальную и максимальную величины признака; здесь эти величины: 29 и 37.
Можно находить среднее арифметическое, пользуясь способом сумм. При этом умножение частот на уклонения заменяется сложением частот. Найдем среднюю длины хвостового стебля воблы ряда и этим способом.
Выпишем частоты нашего ряда, начиная с частоты, соответствующей наибольшей варианте ряда, в виде вертикального столбца. За А мы примем, как и ранее, середину класса 20,5 – 21,5, т. е. 21,05. Для ясности частоту 38, соответствующую нашему А, включим в рамку

Приготовим рядом с нашим столбцом место еще для двух новых столбцов и разместим в этих добавочных столбцах четыре черточки следующим образом: две против частоты 38 в обоих столбцах и две во втором столбце против частот, соседних 38:

Теперь рядом с верхней и нижней частотами напишем их еще раз (в обоих столбцах):

Затем заполняем два новых столбца (т. е. второй и третий) следующим образом: к верхней частоте прибавляем следующую частоту левого столбца и пишем полученную сумму, в данном случае 1+ 6 = 7, в столбце под первой частотой к этой сумме ( к 7) прибавляем следующую частоту (13) и т. д. до черточки. Затем то же делаем и с нижней частью столбца (начинаем снизу: 7 + 9 = 16; 16 +26 = 42). Таким образом заполняем второй столбец:
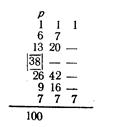
Заполнив второй столбец, таким же образом заполняем и третий столбец, доходя сверху и снизу до черточек (в данном случае приходится находить по одной сумме):
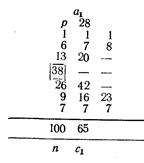
Составив новые столбцы, приступаем к их суммированию; причем сумму верхней части и сумму нижней части второго столбца можно получить, сложив цифры над черточками в верхней части и под черточками-в нижней в обоих новых столбцах. Сумма верхней части 20+8=28(1+7+20=28); сумма нижней части: 42+23=65(42+16+7=65). Эти суммы записаны над столбцом и под столбцом и обозначены а1 и с1. Теперь из суммы верхней части столбца (из 28) вычтем сумму нижней части столбца (65) и получим величину 37, соответствующую ?ра (см. выше). Обозначим эту величину здесь через S1:
S1 = a1- c1 = 28 – 65 = – 37.
Дальнейший ход решения такой же, как и при способе уклонении, а именно:
![]()
Теперь остается отыскать М
М=А + b1 =21,05 + (-0,37) =20,68%.
Получилась величина М такая же, как и при решении способом уклонений. Если решение ряда ограничивается отысканием одной средней М, можно второй новый столбец не заполнять, а суммы верхней и нижней части первого (добавочного) Столбца находить непосредственно из этого столбца.
Средняя арифметическая (М), несомненно, с большей, чем мода, точностью определяет среднюю величину вариационного ряда, т. е. среднее качество признака. Средняя характеризует типичность взятого признака.
В приведенном примере размах варьирования длины хвоста воблы выразился от 18 до 24 (в процентах длины тела), но мы не можем сказать, что эти числа обязательны для любого количества взятых для исследования особей: возьмем меньшее количество рыб – ширина нашего ряда, возможно, сократится, возьмем большее – ряд может расшириться; кроме того, не исключается возможность, что крайние члены этого ряда относятся не к этому, а к другому ряду, т. е. рыбы, которым свойственны крайние величины ряда, входят в другую систематическую группу.
Таким образом, средняя величина не характеризует ни ширину вариационного ряда, ни размах варьирования.
Среднее квадратическое или основное уклонение ?. Нахождение ? способом уклонения и способом сумм
Наилучшей мерой определения размаха или протяженности Вариационного ряда или степени варьирования того или другого Признака в настоящее время считается основное или среднее квадратическое уклонение, обозначаемое буквой ?. (сигма). Для получения этой величины уклонения вариант от средней величины (М), обозначаемые нами буквой а, возводятся в квадрат, перемножаются на их частоты, произведения суммируются, сумма делится на общее число случаев (особей) и из полученного числа извлекается квадратный корень, т. е.
![]()
По этой формуле сигму (о) определяют редко, вычисление среднего квадратического уклонения ведут теми же непрямыми способами, как и среднего арифметического, т. е. по уклонениям от А и способом сумм.
![]() Способ уклонений
Способ уклонений
? = vb2 – b12
Пользуемся тем же примером (стр.301), но здесь уклонения а2 берутся в квадрате: 1, 4, 9,…..
Числа случаев, уклоняющихся и влево от А, здесь должны иметь также знак плюс, потому что отрицательное уклонение (-а), будучи возведенным в квадрат, становится величиною положительной.
Уклонения (а2) 1 4 9
Частота уклонений (р) +26 +9+7
+13 +6 +1
Алгебраическая сумма частот
уклонений (?p) +39 +15 +8
Сумма квадратических уклонений от А, помноженных на частоту
(?pa2): 39 + 60 + 72 = 171,
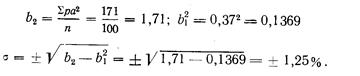
Возведение в квадрат и извлечение корней квадратных можно вести по специальным таблицам.
|
Уклонения в квадрате |
1 |
4 |
9 |
39 |
||
|
+60 |
||||||
|
Простые уклонения |
1 |
2 |
3 |
72 |
||
|
|
– |
26 |
9 |
7 |
||
|
+ |
13 |
6 |
1 |
171 |
100 1,71 b2 = 1.71 |
|
|
Вычислениеb1 |
-13 – 6 – 18 = – 37/ 100 = – 0.37 |
|||||
При пользовании способом уклонений запись вычисленных величин можно упростить следующим образом. Записав уклонения и частоты обычным порядком (причем плюс и минус не нужно повторять, а ставить их один раз слева от частот) и, высчитав b1.записываем над простыми уклонениями уклонения в квадрате. Далее проводим справа вертикальную черту и за чертой записываем в виде столбца произведения сумм частот на уклонения в квадрате, помня, что частоты здесь будут со знаком + (плюс). Данные произведения складываем, делим на п. и получаем значение b2. При такой записи достигается большая экономия времени.
Способ сумм
? = ± ![]()
Пользуемся тем же примером, какой приведен для вычисления средней.
 Ci
Ci
?x2 (в формуле) означает сумму центральных уклонений (т. е. уклонений от M), равную ![]() ; S1 уже найдено (-37), возведем его в квадрат и, разделив на п, получим
; S1 уже найдено (-37), возведем его в квадрат и, разделив на п, получим
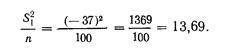
Находим S2 S2 = a1 + с1 + 2(а2 + с2)
а1 и с1 нам известны (a1=28, с1=65; это-суммы верхней и нижней частей второго столбца); а2 – сумма верхней части третьего столбца (8+1=9); с2-сумма нижней части третьего столбца (23+7=30). Таким образом, в числовых выражениях:
S2 = 28 + 65 + 2 (9 + 30) = 93 + 78 = 171.
?x2= 171- 13,69 = 157,31.
Для определения о найденное значение ?x2 делим на n и из полученного частного извлекаем квадратный корень: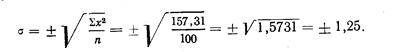
Можно рекомендовать пользоваться при вычислении сигмы комбинированным способом, т. е. соединением обоих способов (способа уклонений и способа сумм). Поясним на примере: Находим S1и S2 (по способу сумм), которые по величине всегда будут равны ?ра и ?ра2. После этого можно перейти к нахождению b1 и b2(по способу уклонений) и, наконец, по формуле ?=± ![]() находим сигму, т. е. среднее квадратическое уклонение:
находим сигму, т. е. среднее квадратическое уклонение:
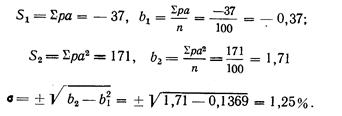
Практическое значение найденных нами величин: М=20,68 и ? =±1,25 таково.
Типичная величина процентного отношения длины хвостового стебля воблы к длине тела равна 20,68%. Чтобы знать, насколько данная форма в этом признаке уклонилась от средней, типичной, формы, нужно знать протяженность вариационного ряда. Если, например, ряд короткий, то и небольшие отклонения от средней будут иметь большее значение, чем такие же отклонения более протяженного ряда. Сигма и показывает протяженность данного вариационного ряда. Чем меньше ?, тем ряд менее изменчив, т. е. протяженность его меньше. Но математически доказано, что 1? не может гарантировать протяженность ряда во всех случаях. Поэтому берут не 1?, а З ? и даже 3,5?.
При нормальном распределении вариант, идеальным случаем которого является распределение коэффициентов двучлена (бинома), возведенного в какую-либо высокую степень, приближающуюся к бесконечности, вариационный ряд и его графическое изображение – нормальная вариационная кривая имеет строго симметричное построение и в таком ряду в промежутке:
от М до ± 1? укладывается около 68% случаев
» М » ±2 ? » » 95,5% »
» М » ±3 ? » » 99,7% »
» М » ±3,5 ? » » 99,95% »
Иначе говоря, более чем на 3? могут уклониться из 100 случаев только три случая. Поэтому, зная одну, две и три сигмы, мы можем определить возможную протяженность нашего конкретного вариационного ряда, следующего закономерности нормального распределения, т. е. можно узнать его крайние варианты. Обратимся к нашему примеру.
Класс: 17,5-18,5-19,5-20,5-21,5-22,5-23,5-24,5 n
Частота 7 9 26 38 13 6 1 100
M=20,68 ?=±l,25 3 ?=±3,75
Следовательно, крайние члены ряда, если основываться на 3?, должны быть 16,93 (20,68-3,75) и 24,43 (20,68+3,75), т. е. наш первый эмпирический ряд меньше второго теоретического, а это говорит о том, что мы можем встретить воблу, длина хвостового стебля которой может быть меньше той величины, какую мы наблюдали, т. е. не 18, а 17%, но правая сторона ряда и теоретически не превосходит 24%, т. е. с более длинным хвостовым стеблем воблу едва ли встретим. Для большей осторожности лучше брать не 3, а 3,5?. Рассчитав протяженность ряда по 3,5?, получим 16,30-25,06; при таком расчислении можно предполагать три новых варианта (16, 17 и 25%).
При небольшом числе наблюдений (менее 25) основное уклонение следует определять по формуле
![]()
где сумма квадратических уклонений от М, умноженных на частоту, делится на количество наблюдений без единицы (n-1).
В этой формуле выражение ?p?2 не может приниматься тождественным выражению ?ра2, где а2 есть уклонение от А (от приближенной средней).
Отыщем ? при n =15:
Число жаберных тычинок 29 30 31 32 33 34 35 36 37 n
Частота 1 1 2 5 2 2 0 0 2 15
Так как здесь принимаются отклонения от М, то необходимо сначала определить М. М=32,60.
Находим уклонения от нашего М, т. е. от 32,60.

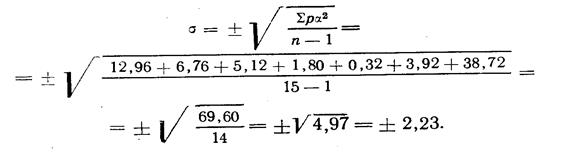
Если бы вычисления шли при неуменьшенном n, т. е. при 15, ii не при 14 (n-1), то ? равнялась бы ±2,15. Различие сигм для малого п (15) существенно.
При работах по определению морфологических признаков рыб вообще не следует вести вариационно-статистическую обработку при малом количестве особей (менее 25) ,апотому и пользоваться рассмотренной формулой (![]() ) почти
) почти
не приходится. Если же приходится, то во всяком случае не следует вычисления сигмы вести таким длительным способом, какой мы только что рассмотрели, а решать способом уклонений или способом сумм.
К решению формулы ![]() можно придти, пользуясь формулой
можно придти, пользуясь формулой
![]()
Решение ряда можно вести так

Ход решения данного примера можно вести и иначе. Составляем решетку.
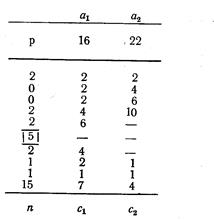
Решение идет таким образом
![]()
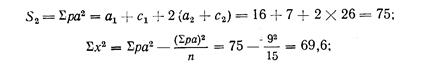
отсюда
![]()
В рядах,где классовый промежуток не единица, найденное значение сигмы нужно помножить на величину классового промежутка (?):
![]()
Коэффициент вариации С и средние ошибки М, ? и С
![]()
Сигма определяет степень варьирования признака, степень изменчивости вариационного ряда, но при сравнении двух признаков, у которых М различно, сравнение сигм не создает правильного представления об изменчивости данных признаков. Предположим, что в одном случае ? =±1,25, М=20,68; в другом случае ?=±2, а М=38. Какой признак наиболее изменчив?
Если бы средние были приблизительно одинаковы, мы могли бы сказать, что ряд с ? = ±2 более протяженный и значит более изменчивый. Но при различных средних может оказаться, что ряд с сигмой, равной ±1,25, более изменчив.
Чтобы сравнить изменчивость этих двух рядов, нужно сигму выразить в процентном отношении к средней величине того же ряда, к которому относится сигма. Это процентное отношение ? к М и называется коэффициентом вариации. Коэффициент вариации обозначаем буквой С.
В нашем примере ? = 1,25.
![]()
В другом случае (при ?=±2):
![]()
Таким образом, ряд с большой сигмой оказался менее изменчив, чем ряд с сигмой, равной ±1,25. При помощи С мы можем сравнивать любые признаки, например вес рыбы и ее размер, выраженные в любых единицах измерения, так как С есть величина отвлеченная (процент от М). Таким образом, коэффициент вариации является мерилом относительной степени изменчивости определенного признака.
Средние ошибки М, ? и С
Необходимо указать на способы нахождения средних ошибок: средней величины (М), основного уклонения (?), коэффициента вариации (С) и разности средних величин (M1-М2). Формулы ошибок первых трех величин очень просты и не требуют никаких пояснений.
Средняя ошибка М или ![]() В тексте среднюю ошибку можно обозначить ±m и ставить рядом с той величиной, к которой она относится: М±т, ?±т (для краткости пишут Мm, ?т и т. д.).
В тексте среднюю ошибку можно обозначить ±m и ставить рядом с той величиной, к которой она относится: М±т, ?±т (для краткости пишут Мm, ?т и т. д.).
Средняя ошибка показывает возможное колебание средней М. В нашем примере средняя величина получилась равной 20,68%. В другом случае при подобных же измерениях длины хвостового стебля воблы М может получиться иное. Предел колебания средней и определяется формулой
![]()
В данном примере
![]()
или, округляя до сотых, тм= ±0,13,
т. е. М может быть более или менее 20,68 на 0,13.
Ошибка средней, как видно из формулы, зависит от количества наблюдений: чем больше число измеренных особей, тем ошибка меньше; она обратно пропорциональна корню квадрат ному из числа измерений. Значит, от нее зависит величина ошибки. Полученная нами величина вместе с ее средней ошибкой должна быть записана так: М ± m= 20,68 ± 0,125
или короче
Мm=20,68 ±0,125.
Средняя ошибка квадратического уклонения находится по формуле
![]()
В наших работах этой ошибке обычно придается малое значение, равно как и ошибке коэффициента вариации, которая высчитывается по формуле
![]()
Значение ошибки квадратического уклонения и ошибки коэффициента вариации то же, что и ошибки М, т. е. т определяет колебание величин, к которым ошибка относится.
Дифференция рядов
![]()
Наибольшее значение при ихтиологических работах имеет средняя ошибка разности средних величин признаков двух групп рыб. При установлении вида, подвида, расы и морфы необходимо пользоваться формулой дифференции или расхождения рядов. Многие авторы в своих статьях называют эту формулу diff. (сокращенное слово differentia):
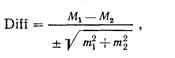
где M1-М2–разность средних величин взятых рядов, а ![]() -средняя ошибка этой разности. Из выражения
-средняя ошибка этой разности. Из выражения ![]() видим, что средняя ошибка разности средних величин равна квадратному корню из суммы квадратов средних ошибок, соответствующих данным средним величинам: т21 – квадрат средней ошибки M1, a m22 – квадрат средней ошибки M2.
видим, что средняя ошибка разности средних величин равна квадратному корню из суммы квадратов средних ошибок, соответствующих данным средним величинам: т21 – квадрат средней ошибки M1, a m22 – квадрат средней ошибки M2.
Возьмем два вариационных ряда, определяющих отношение длины хвостового стебля к телу рыбы для воблы и для формы, близкой к вобле,- серушки (Rutilus rutilus fluviatilis).
16,5-17,5-18,5-19,5-20,5-21,5-22,5-23,5-24,5 n
– 7 9 26 38 13 6 1 100
7 31 30 17 11 3 – – 99
Перед нами две формы, которые по длине хвостового стебля резко не различаются, потому что многие варианты свойственны и той и другой группе (пример трансгрессивных рядов).
Хотя в крайних величинах расхождение и замечается; но числа, свойственные только одному ряду, очень малы: в первом ряду только 7 рыб из 100 можно бесспорно отнести к R. rutilus caspicus, а остальные 93 по этому признаку можно относить и к R. rutilus caspicus и к R. rutilus fluviatilis. Для таких случаев биометрика и дает формулу
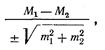
которая обозначает отношение разности средних к средней ошибке этой разности, причем на основании теории вероятностей принято считать, что если эта разность средних больше ее средней ошибки в три раза и более, то такая разность делает различие вариационных рядов сильным, или, по довольно распространенному, но совсем не точному выражению, — реальным.
![]() Чтобы не извлекать квадратного корня из можно разницу M1-М2 возвести в квадрат. Это иногда бывает проще выполнить, причем и число 3 (прежний показатель разницы рядов) нужно возвести так же в квадрат; тогда новый показатель разницы должен быть равным 9.
Чтобы не извлекать квадратного корня из можно разницу M1-М2 возвести в квадрат. Это иногда бывает проще выполнить, причем и число 3 (прежний показатель разницы рядов) нужно возвести так же в квадрат; тогда новый показатель разницы должен быть равным 9.
При работах по систематике рыб, если исследователь не учтет зависимости между признаками (см. ниже о корреляции) или влияющих на изменение данного признака биологических факторов, то один математический расчет по формуле:
![]()
не дает права говорить о двух различных рядах.
Возвратимся к примеру.
Для первого ряда нам известны: средняя, основное уклонение и ошибка средней:
М = 20,68
? =±1,25
mМ=±0,12
Находим эти величины и для второго ряда, руководствуясь приемами, рассмотренными выше.
Способ уклонений
![]() За условную среднюю (А) берем 19,05. Отклонения ( от А)
За условную среднюю (А) берем 19,05. Отклонения ( от А)
![]() Частота уклонений
Частота уклонений
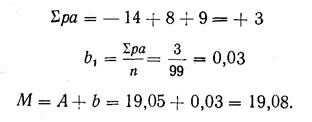
Определяем основное уклонение (?):
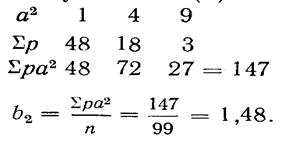
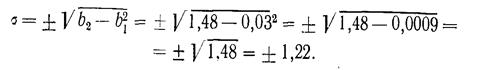
Способ сумм (комбинированный)
 Средняя ошибка средней величины второго ряда
Средняя ошибка средней величины второго ряда
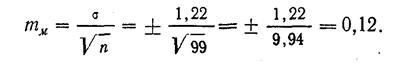
Отыскиваем разность средних, среднюю ошибку этой разности и определяем отношение между этой разностью и средней ошибкой:
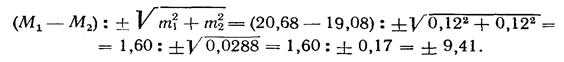
![]() Если пользоваться упомянутым выше способом, исключающим извлечение квадратного корня из то решение пойдет так
Если пользоваться упомянутым выше способом, исключающим извлечение квадратного корня из то решение пойдет так
![]()
![]() При первом решении (при извлечении квадратного корня из ) показатель отношения разности средних к ее ошибке выразился числом, близким к 9 (1,60:0,17=9,41); во втором случае такой же показатель выразился числом 88,89, т. е. близким к квадрату 9.
При первом решении (при извлечении квадратного корня из ) показатель отношения разности средних к ее ошибке выразился числом, близким к 9 (1,60:0,17=9,41); во втором случае такой же показатель выразился числом 88,89, т. е. близким к квадрату 9.
Приведенная разница показывает, что взятый признак (длина хвостового стебля) является хорошим отличительным признаком одной формы плотвы – Rut. rut. caspicus от другой Rut. rut. fluviatilis.
Когда ведут сравнение признака сразу у нескольких форм, полезно пользоваться простым и наглядным описанным мною приемом табличного выражения результатов анализа формулы (Правдин, 1925):
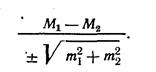
Пример. Сравниваем число чешуи в боковой линии пяти форм плотвы Rut. rut. typicus, Rut. rut. fluviatilis, Rut. rut. caspicus, Rut. rut. var. ? и Rut. rut. var. ? (табл. 17).
В клетках таблицы указаны частные от деления разности числа чешуи в боковой линии взятых форм на ее ошибку.
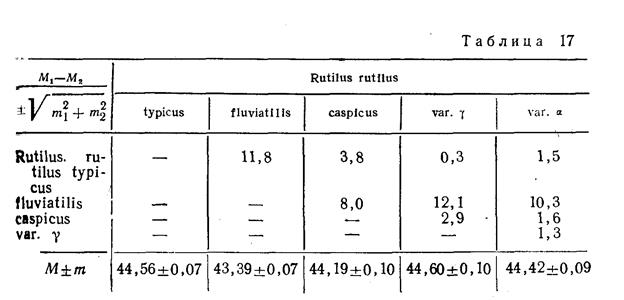
Посмотрим, есть ли различие в числе чешуи Rut. rut. typicus и Rut. rut. caspicus. Клетка в пересечении горизонтальной графы с надписью Rut. rut. typicus с графой вертикальной, где подписано Rut. rut. caspicus, содержит число 3,8, показывающее, что разность в числе чешуи взятых форм превосходит ошибку более чем в 3 раза, т. е. разность существенна. Напомним, что число чешуи в боковой линии можно подсчитать безошибочно и что этот признак почти не зависит от возраста и пола плотвы, поэтому число 3 достаточно для того, чтобы различие считать -значительным. Между типичной плотвой и плотвой у различий и этом признаке нет (cliff =0,3); нет различия и между Rut. rut. var. ? и Rut. rut. var. ? (diff =1,3) и т. д. Наша таблица выражает не только расхождение признаков, но и схождение, родственность их. Rut. rut. fluviatilis по числу чешуи в боковой линии дальше всего отстоит от Rut. rut. var. ? (diff.= 12,1), затем от Rut. rut. typicus (diff. = 11,8), потом от Rut. rut. var. ? (diff.= 10,3) и, наконец, от Rut. rut. caspicus (diff.=8), т. е. Rut. rut. fluviatilis ближе стоит по числу чешуи в ll к Rut. rut. caspicus-форме, к которой Rut. rut. fluviatilis близка и по своему территориальному распространению. Нижняя горизонтальная графа, содержащая показатель средних величин числа чешуи в ll каждой взятой формы, дает возможность быстро узнавать, у какой формы число чешуи больше, у какой меньше. Подобными таблицами можно изобразить любые признаки любых форм.
А. В. Морозов предложил простой способ нахождения различия между отдельными пробами. Метод этот состоит в вычислении суммы различий, определения их средних (М diff.) и D сравнении их по отдельным пробам. Наглядно такой метод может быть представлен в таблице, взятой из работы Е. А. Безруковой (1939) (табл. 18).
Взяты были четыре пробы подустов (донского, волжского, камского и днепровского) по четырем счетным признакам. Найдены соответствующие расхождения (diff.) между каждой парой проб в каждом признаке, затем установлена сумма расхождении каждой пары и среднее отличие.
Таблица 18
Степень реальности различий между отдельными пробами по меристическим признакам (по Безруковой, 1939)
|
Название признака |
Дон Волга |
Дон Кама |
Дон Днепр |
Волга Кама |
Волга Днепр |
Днепр Кама |
|
Чешуи в И ………….…………….. |
5,02 |
10,70 |
13,12 |
3,79 |
5,95 |
2,53 |
|
Лучей в D …………………………. |
1,00 |
0,89 |
2,44 |
1,40 |
1,05 |
2,46 |
|
Лучей в Л ………….……………… |
0,20 |
2,40 |
2,93 |
2,28 |
2,50 |
4,53 |
|
Количество позвонков………. |
11,76 |
6,38 |
20,94 |
0,20 |
9,18 |
5,33 |
|
Сумма отличий………………….. |
17,96 |
20,37 |
39,43 |
7,67 |
18,68 |
14,85 |
|
Среднее отличие (Мdiff) ……. |
4,50 |
5,09 |
9,86 |
1,92 |
4,67 |
3,71 |
В результате можно говорить о существенном расхождении в меристических признаках между подустом донским и камским (Мdiff =5,09) и об отсутствии расхождения между волжским и камским подустами (Мdiff = 1,92). Точно таким же способом можно вести сравнение проб и по пластическим признакам.
Таким образом, мы элементарно рассмотрели самые существенные случаи и способы применения вариационной статистики к работам ихтиологической систематики. Остановимся также на применении вариационной статистики к случаям, когда необходимо выяснить зависимость одного признака от другого, к случаям коррелятивной зависимости.
В этой книге мы уже говорили, что многие признаки рыб находятся между собой в прямой зависимости: при увеличении (при росте) рыбы в длину высота тела и антедорсальное расстояние увеличиваются не только в абсолютных значениях, но и в относительных. Но величина глаза, длина головы и некоторые другие части тела при росте рыбы в длину, мало увеличиваясь в абсолютных значениях, в относительных-уменьшаются, поэтому мы и говорим, что у взрослой рыбы голова относительно короче, глаз меньше. Представим, что мы имеем две группы одного и того же вида рыбы: одна группа состоит из более молодых, другая – из более взрослых рыб. Положим, мы подметили, что глаза у рыб первой группы крупнее, а тело выше, чем у второй. Можем ли мы сделать вывод, что перед нами две породы, две расы, или что одна группа рыб состоит из типичных представителей данного вида, а другая является расой этого вида? Нет, не можем, размер глаза и высота тела рыбы зависят от ее возраста. Поэтому необходимо знать, во-первых, действительно ли у взятых групп эта зависимость есть, и во-вторых, в каком направлении и в какой мере эта зависимость выражается и изменяется. На эти вопросы дают ответы коэффициент корреляции и коэффициент регрессии. Та и другая величины имеют свое научное теоретическое обоснование, но мы здесь ограничимся короткими замечаниями и дадим лишь технические приемы для вычисления названных -коэффициентов.
Коэффициент корреляции
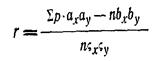
Различают корреляцию положительную, когда с увеличением одного признака, одного свойства, увеличивается и другой признак, и отрицательную, когда с увеличением первого свойства уменьшается второе.
Пример. Желательно выяснить, есть ли корреляция между величиной глаза плотвы, описанной нами под названием Rut. rut. var. ? и длиной ее тела. Для этого необходимо составить так называемую решетку или таблицу корреляции. Материалом для этой таблицы нам послужат два вариационных ряда: классовые вариации длины тела (в мм) и такие же вариации размера глаза (в % длины тела).
Длина тела, мм
81-100 101-120 121-140 141-160 161-180 181-200 201-220 221-240
16 11 5 12 10 5 2 3
241-260 261-280 281-300 п
1 2 3 70
Диаметр глаза (в % длины тела)
3,5 – 4,5 – 5,5 – 6,5 – 7,5 – 8,5 п
9 24 17 17 3 70
Эти два ряда в таком виде еще не говорят об их взаимной зависимости; удобнее изобразить эти ряды на сторонах прямоугольной сетки: на одной стороне сетки разместим классы длины тела, на другой наружной стороне – классы диаметра глаза, а в отдельных внутренних клетках этой решетки поставим, сколько особей с данным диаметром глаза относится к каждому классу длины тела (табл. 19).
Таблица 19
|
Диаметр глаза (в % длины тела |
Длина тела, мм |
|||||||||||
|
81-100 |
101-120 |
121-140 |
141-160 |
161-180 |
181-200 |
201-220 |
221-240 |
241-260 |
261-280 |
281-300 |
? |
|
|
4 5 6 7 8 |
– – 1 12 3 |
– – 6 5 – |
– – 5 – – |
– 8 4 – – |
– 9 1 – – |
– 5 – – – |
1 1 – – – |
2 1 – – – |
1 – – – – |
2 – – – – |
3 – – – – |
9 24 17 17 3 70 |
|
? |
16 |
11 |
5 |
12 |
10 |
5 |
2 |
3 |
1 |
2 |
3 |
|
Эта таблица наглядно показывает, что с увеличением длины тела рыбы величина глаза относительно падает-пример отрицательной корреляции.
Возьмем другой пример. Зависимость высоты тела рыбы от ее роста в длину. Корреляционная таблица этих рядов признаков будет иметь другой характер (табл. 20). Заметим, что свойство, от которого зависит другое, называется подлежащим и обозначается через х, а зависимое свойство обозначается у. Выбор из двух свойств одного
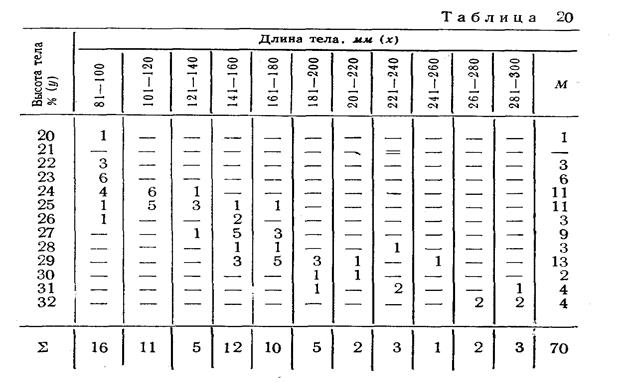
Из таблицы ясно видно, что с увеличением длины рыбы высота ее тоже увеличивается, т. е. это положительная корреляция.
Корреляционные таблицы наглядны, если они представляют: одинаковое число (и не менее 6-8) классов того и другого признака, т. е. когда корреляционная сетка близка к форме квадрата и когда число взятых особей рыб более 50 (лучше 100).
В обоих приведенных примерах корреляция есть, но какова степень корреляционной зависимости? Для этого надо найти коэффициент корреляции, который обозначается буквой г и находится по формуле
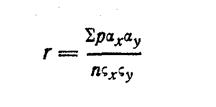
или (чаще)
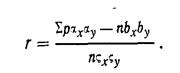
Объяснение этих формул можно найти в книге проф. Ю. А. Филипченко «Изменчивость и методы ее изучения» (1929) или в любом из новейших учебников по вариационной статистике (биометрике), список которых приведен ниже. Ограничимся подробным решением конкретного примера корреляционной таблицы, приведенной нами для выяснения зависимости между длиной рыбы и диаметром глаза.
Вычисления для большей простоты нужно вести так же, как и при отыскании средней величины (М) и основного уклонения ?. Удобнее пользоваться способом уклонений от приближенной средней, причем классовый промежуток в расчет не принимается, для чего здесь пишется не обычная сигма (?), а сигма круглая (?).
В обоих рядах какой-либо класс (лучше средний) принимается за приближенную среднюю Л, и от нее ведутся все вычисления. Этот класс будет иметь нулевое уклонение п, и обозначается нулем (0). В ряду классов длины тела за А принимается класс 181-200, в ряду классов диаметра глаза – класс 6, вернее 5,6-6,5.
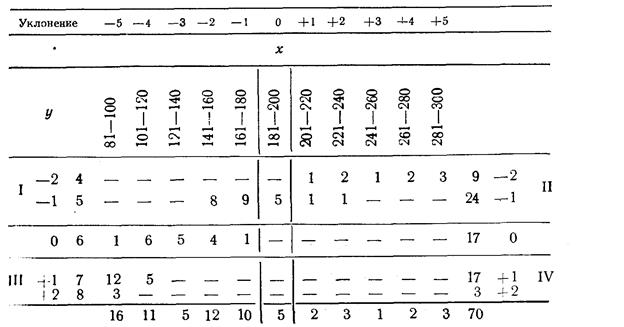
Пересекающиеся нулевые классы разбивают таблицу на четыре части (на четыре квадранта): I, II (вверху) и III и IV (внизу). В каждой клетке нужно частоту перемножить на соответствующие уклонения в том и другом ряду и для каждого квадранта найти сумму. Произведения в I и IV квадрантах получатся со знаком плюс, так как частота в каждой клетке I квадранта умножается два раза на отрицательную величину, а произведение величин с одинаковыми знаками дает плюс (в IV квадранте перемножаются положительные уклонения); произведения II и III квадрантов по такой же причине будут со знаком минус, так как в них частоты перемножаются на одно положительное и одно отрицательное уклонение. Решение примера удобнее вести так:

Общая сумма произведений уклонений на частоты во всех четырех квадрантах
![]()
Определяем b и ? в каждом ряду, значение b отождествим с известным уже нам значением b(bх и by), a ? с ?, т. е. определяем простое и квадратическое уклонения от А в ряду х и в ряду у.
Получим для первого х ряда bх:
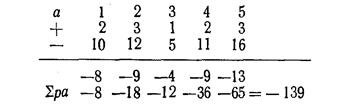
![]() Таким же путем находим by для второго (у) ряда:
Таким же путем находим by для второго (у) ряда: ![]()
?xопределяем как ? первого ряда.
?x=±2,76, а ?у -как ? второго ряда. ?x =±l,0.
Подробное решение приведенных рядов (х и у) можно вести тем же порядком, который указан выше по способу уклонений, а здесь дадим подробное решение тех же рядов, пользуясь комбинированным способом (решетки рядов х и у см. ниже).
Решение ряда х
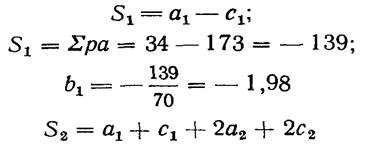
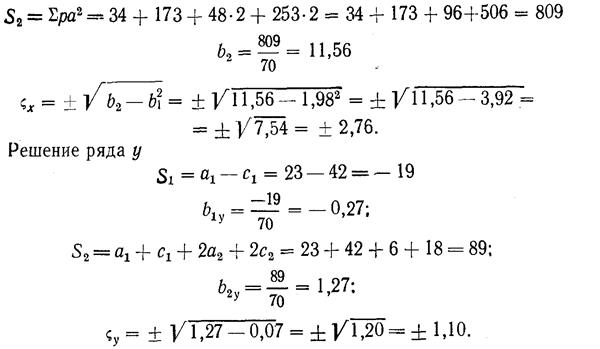
Заменяя буквы формулы их найденными значениями, получим

Коэффициент корреляции
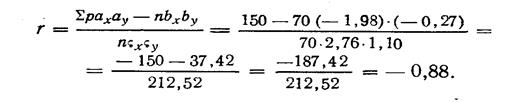
Между рядами, т. е. между длиной тела плотвы и диаметром ее глаза, очень высокая отрицательная корреляция, которая выражается коэффициентом 88%. Когда корреляция имеет коэффициент около 50%, то она считается средней. Корреляция с коэффициентом 70-80% считается очень высокой, а с коэффициентом 20-30% -слабой.
Средняя ошибка коэффициента корреляции находится по формуле:
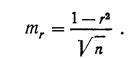
При ихтиологических работах средней ошибкой корреляционного коэффициента пользоваться нужно.
Коэффициент корреляции указывает на зависимость между двумя признаками или двумя свойствами, но не может ответить на вопрос: в каком размере происходит изменение одного признака в зависимости от другого.
На это отвечает коэффициент регрессии, причем регрессией называется соотношение изменения одного свойства с изменением другого.
Коэффициент регрессии R
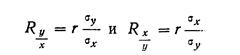
![]()
![]() Коэффициент регрессии обозначается буквой R,причем, если нас интересует изменение второго (зависимого) свойства в связи с изменением первого (подлежащего), то это обозначается если же, наоборот, узнается изменение первого в связи с изменением второго, то обозначается Первый
Коэффициент регрессии обозначается буквой R,причем, если нас интересует изменение второго (зависимого) свойства в связи с изменением первого (подлежащего), то это обозначается если же, наоборот, узнается изменение первого в связи с изменением второго, то обозначается Первый
![]()
коэффициент определяется по формуле второй-по формуле
![]()
При вычислении сигмы здесь учитываются величины классовых промежутков (поэтому пишется сигма конечная, а не круглая).
Предположим, что желая выяснить коэффициент регрессии между возрастом и длиной тела сигов, найдем:
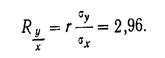
Это значит, что при увеличении возраста рыбы на 1 год длина ее увеличивается на 2,96 см; при увеличении же на 2 года можно ожидать увеличения длины тела на 5,92 см. Если бы мы решили ![]() , то этот же результат показал бы нам, что при увеличении длины рыбы на 2,96 см нужно ожидать, что ее возраст увеличился на один год.
, то этот же результат показал бы нам, что при увеличении длины рыбы на 2,96 см нужно ожидать, что ее возраст увеличился на один год.
Средняя ошибка коэффициента регрессии вычисляется по формулам
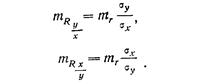
Уделяя исключительно большое внимание методам биометрики и считая совершенно немыслимой работу современного ихтиолога без знакомства с основами этих методов и без введения в ихтиологическую работу элементов вариационной статистики, автор все же предостерегает от излишнего увлечения математическими выкладками. Чисто механическое пользование коэффициентом регрессии в таких работах, как систематика рыб, может привести к неверным выводам, так как темп роста ‘тела рыбы в длину изменяется с возрастом, в молодом возрасте рыба растет быстрее, чем в старшем. Поэтому коэффициент регрессии длины и возраста для более старых рыб будет меньше, чем у рыб более молодых.
Другие методы, например метод обратного расчисления длины рыбы по данной величине чешуи, несмотря на его некоторую незаконченность, даст в подобном случае результат более надежный, чем коэффициент регрессии.
Вариационная статистика, учитывая (в числах) те или иные признаки и свойства организмов, почти не касается биологической природы наблюдаемых явлений. Между тем изучение такого объекта, как рыба, представляет много своеобразных биологических особенностей, которые влияют не только на внутреннее строение рыбы, но и на ее внешние признаки. Мы не раз указывали на особенности роста рыбы в длину, на особенность роста головы и т. п. Все это обязывает нас вести строгий учет биологических явлений, свойственных изучаемым нами рыбам.
Общие замечания к вариационно-статистическим работам
1. Для промеров брать около 100 экз. (лучше 100) взрослых рыб разного размера, разного пола и различной степени зрелости. Рыб с явно выраженными сезонными изменениями для целей систематики брать не следует. Самцов и самок следует рассмотреть отдельно и, если будет заметно морфологическое расхождение полов, обработать отдельно материал по самцам и отдельно по самкам. Число 100 взято на основании опыта автора по обработке огромного материала по плотве и сигам, но оговоримся, что бывают случаи, когда можно взять рыб и меньше 100 особей (это бывает, когда исследуемая группа заведомо не обнаруживает склонности к морфологическому варьированию). Количество больше 100 часто совершенно не дополняет и не изменяет вариационного ряда, составленного на основании измерения 100 рыб. Число 100, кроме того, облегчает решение многих формул, ибо на 100 делить, умножать или извлекать корень из 100 легко. Пользуясь указаниями теории вероятности, можно сделать расчет нужного для избранной точности количества наблюдений, но все это в большинстве случаев будет расчетом для наших работ далеко не бесспорным и не легким.
В руководствах по вариационной статистике указываются формулы для определения нужных количеств наблюдений, но в наших работах лучше пользоваться тем общим положением, что средняя ошибка должна быть меньше той величины, к которой она относится, не менее чем в три раза. Но в действительности ошибка средних, получаемых при определении морфологических признаков рыб, всегда бывает меньше самой средней во много раз (до 100 раз).
2. Брать для промеров возможно большее количество признаков, помня, что такой материал весьма ценен для многих вопросов все еще весьма мало разработанной теории систематики рыб.
3. Для выяснения корреляционной зависимости необходимо брать ряды не менее чем с 6-8 классами и суммой наблюдений не менее 50.
4. Пределы теоретического вариационного ряда лучше высчитывать 3,5 сигмами.
5. Различие между вариационными рядами, т. е. дифференцию (diff.), считать существующей тогда, когда формула при решении дает частное больше трех.
![]()
6. Методом Гейнке, т. е. методом комбинированных признаков, пользоваться нужно редко, потому что отнесение любой особи к ее типу (это имеет место чаще всего при работах над расами рыб) возможно лишь тогда, когда тип имеет подробнейшее описание, точную характеристику многочисленных признаков. Это обстоятельство и удержало меня от изложения метода Гейнке.
7. При работах над расами рыб морфологические признаки и отличия необходимо рассматривать параллельно с признаками биологическими. Материал для изучения расового состава рыб того или иного водоема надо собирать в период нереста.
В случаях, когда половой диморфизм не выражен, можно не разделять материал по полу. Прежде чем приступить к выяснению различия между расами по тем или иным признакам, необходимо установить наличие изменчивости этих признаков в зависимости от длины тела и возраста рыб. При обработке расового материала необходимо обращать внимание на темп роста и на форму и структуру чешуи. При определении дифференции между признаками необходимо учитывать половые и возрастные различия.
8. При описании группы рыб надо давать сводную ведомость, в которой указывать М±т, а, С и крайние варианты теоретического вариационного ряда, вычисляемого по 3,5 а (тут же нужно указывать и крайние варианты эмпирического ряда).
9. Желательно при описании рыб опубликовывать подробные ведомости индексов и промеров.
10. Метод вариационной статистики в области ихтиологических исследований применяется не только в работах по систематике рыб, не менее существенную пользу оказывает он и в работах по биологии рыб (особенно при работах по темпу роста). Нужно пользоваться этим методом и ,при изучении промысла – вообще всегда, когда имеется цифровой (количественный или качественный, счетный или измерительный) материал.
Приемы вариационной статистики освещены в новейших руководствах и учебниках, список которых прилагается:
Бейли Н. Статистические методы в биологии. Изд. Иностранной литературы. М., 1962.
Варден Б.Л. Математическая статистика. Изд. Иностранной литературы. М., 1960.
Плохинский Н.Л. Биометрия. Изд. Сибирского отделения АН СССР. Новосибирск, 1961.
Рокицкий П.Ф. Основы вариационной статистики для биологов. Изд. Белгосуниверситета им. В. И. Ленина. Минск, 1961.
Романовский В.И. Применения математической статистики в опытном деле. ОГИЗ. Гостехиздат. М.-Л., 1947.
Снедекор Дж.У. Статистические методы в применении к исследованиям в сельском хозяйстве и биологии. Изд. с.-х. литературы, журналов и плакатов. М.,1961.
Фишер Р. А. Статистические методы для исследователей. Госстатиздат. М, 1958.
Урбах В. Ю. Математическая статистика для биологов и методикой. 11.1Д. ЛИ СССР. М., 1963.
Литература
Абдурахманов Ю.А. Рыбы пресных вод Азербайджана. Баку. Изд. АН Азерб. ССР, 1962.
Абрамов В. В. Mugil cephalus (Linne) Cuvier-лобан из низовья Амура. Доклады АН СССР. Т. 85, № 2, 1952.
Аверинцев С. В. Материалы к познанию промысловых рыб и рыболовства Баренцева моря в связи с проектами дальнейших исследований. Труды Научного института рыбного хозяйства. Т. 2, Вып. 3, 1927.
Аверинцев С. В. К вопросу об изучении рас у рыб. Бюллетень рыбного хозяйства, 1929, № 6.
Аверинцев С. В. Материалы по биологической статистике промысловых рыб в низовьях Лены за 1930 г. Труды Якутской научной рыбохозяйственной станции. Т. 2, 1933.
Аверинцев С. В. Определение промыслового запаса и методы долгосрочных прогнозов в морском рыболовстве. М., 1948.
Аврех В. В. и Калабухов Н.И. Кровное родство горных и равнинных форм лесной мыши (Apodemas sulvaticus) и других близких видов мышей. Зоологический журнал. Т. 16. Вып. I, 1937.
Аврутина Э. Методика определения возраста и роста осетровых рыб по лучу грудного плавника. «Рыбное хозяйство», 1938, № 11.
Алеев В. Р. Угловой или переменный масштаб в применении к обратному расчислению роста рыб по чешуе. «Рыбное хозяйство», 1937, № 4.
Алеев Ю. Г. Функциональные основы внешнего строения рыбы. М., Изд-во АН СССР, 1963.
Александров А. И. Годовой отчет о работах Керченской ихтиологической лаборатории за 1924 г. Труды Керченской ихтиологической лаборатории. Т, I. Вып. I, 1926.
Александрова К. Миграция на кефаловите Риби по нашето краебе режие. «Рибно стопанство», 1957, № 6-7.
Алтухов Ю. П. О влиянии пола и возраста некоторых рыб на теплоустойчивость и степень серологической близости их мышечных белков. «Цитология», 1961, Т. 3, №5.
Алтухов Ю. П. и Апекин B.C. Серологический анализ родственных взаимоотношений «крупной» и «мелкой» ставриды Черного моря. «Вопросы ихтиологии». Т. 3. Вып. 26, 1963.
Амброз А. И. Сельдь (Clupea harengus pallasi С. V.) залива Петра Великого. Биологический очерк. Известия ТИНРО. Т. 6., Владивосток, 1931.
Амброз А. И. Рыбы Днепра, Южного Буга и Днепровско-Бугского лимана. Киев, изд. АН УССР, 1956.
Амирханов Г. А. Иммуно-серологическая характеристика осетровых рыб Каспия. Сб. научно-технической информации ВНИРО. Вып. I, 1964 а.
Амирханов Г. А. Изучение внутривидовых особенностей осетровых рыб Каспия при помощи комплекса иммуно-серологических реакций. Труды молодых ученых. Изд. ВНИРО, 1964 б.
Амосов В. А. О новом показателе упитанности рыб – индексе удельной вальковатости. «Вопросы ихтиологии». Вып. 17, 1961.
Амосов В. А. О форме тела и мясистости пеляди Coregonus peled (Gmelin) озера Врево. «Вопросы ихтиологии». Т. 3. Вып. I, 1963.
Андреев В. Г. Повышение уловистости речных закидных неводов. «Рыбное хозяйство», 1949, № 1.
Андрияшев А. П. Рыбы Северных морей СССР. Изд. АН СССР, 1954.
Арнольд И.Н. К вопросу об определении возраста рыб. «Вестник рыбопромышленности», 1911, № 5 и 6.
Асланова Н.Е. Мечение промысловых рыб. «Вопросы ихтиологии». Т. I. Вып. 3 (20), 1961.
Астанин Л. П. Об определении возраста рыб по костям. Зоологический журнал. Т. 26. Вып. 3, 1947.
Атлас Ленинградской области и Карельской АССР. Л. 1935.
Атлас океанографических основ рыбопоисковой карты Южного Сахалина и южных Курильских островов. 1955, 1956, Изд. ЗИН АН СССР и ТИНРО.
Атлас промысловых рыб СССР. М., Пищепромиздат, 1949.
Базикалова А.Я., Каллиникова Т. П., Михин В.С. и Талиев Д. Н. Материалы к познанию бычков Байкала. Труды Байкальской лимнологической станции. Т. 7, 1937.
Балагурова М. В. Биологические основы организации рационального рыбного хозяйства на Сямозерской группе озер Карельской АССР. Изд. АН СССР, 1963.
Балахнин И. А. Некоторые данные о внутривидовых различиях крови форелей и методика их определения. Доклады АН СССР. Т. 141, № 2, 1961.
Балахнин И. А. Физиологические методы изучения родственных связей рыб. Сборник научно-технической информации, № 7, ВНИРО, 1962.
Баранов Ф.И.К вопросу о биологических основаниях рыбного хозяйства. Известия Отдела рыбоводства и научно-промысловых исследований. Т. I. Вып. I, 1918.
Баранов Ф.И.К вопросу о динамике рыбного промысла. Бюллетень рыбного хозяйства, № 8, 1925.
Барач Г. П. Рыбы пресных вод Абхазии. Материалы по фауне Абхазии. Изд. Грузинского филиала АН СССР, 1939.
Барач Г. П. Рыбы Армении. Труды Севанской гидробиологической станции. Т. 6, 1940.
Барач Г. П. Фауна Грузии. Т. I. «Рыбы пресных вод». Изд. АН Груз. ССР, 1941.
Барсуков В. В. О гидродинамических качествах хвостового плавника атлантических зубаток (Anarhichadidae). Доклады АН СССР. Т. 129, № 3, 1959 а.
Барсуков В. В. Эволюция электрических органов у рыб. «Природа», 19596, № 11.
Барсуков В. В., Иосида И. Новый способ фиксации рыб с сохранением естественной окраски. 1962, Киото (рецензия). Зоологический журнал. Т. 42. Вып. 12, 1963.
Безрукова Е.А. Подуст (Chondrostomanasus) p. Дон. Бюллетень биологической станции Воронежского государственного педагогического института. Т. I. Вып. I, 1939.
Бельчук Д. А. К вопросу об определении возраста одноперого терпуга (Pleurogrammus azonus). Известия ТИНРО. Т. 14, 1938.
Беляев В. Н. Осетр. Asipenser guldenstadti Brandt. Бюллетень Всекаспийской рыбохозяйственной экспедиции, № 5-6, Баку, 1932.
Беляева К. И. Судак (Онежского озера). В кн. «Озера Карелии». Петрозаводск. Гос. изд-во Карельской АССР, 1959.
Белый Н. Д. Изучение нереста и получение икры судака при помощи искусственных гнезд. II. Экологическая конференция. Тезисы докладов. Т. 3. Изд. Киевского гос. университета, 1951.
Білий Н. Д. Нерест дніпровського судак та здобування його ікри за допомогою штучних гнізд. Труди Інст. гідробіології АН УССР, № 27, 1952.
Белый Н. Д. Размножение тарани Rutilus rutilus heckeli (Nordmann) в низовьях Днепра на искусственных гнездах. «Вопросы ихтиологии». Т 1 Вып. 3 (20), 1961 а.
Белый Н. Д. Размножение рыб на искусственных гнездах в Каховском водохранилище. «Рыбное хозяйство», 1961 б, № 6.
Бенинг А. Л. О плодовитости стерляди. Известия Института сельского к хозяйства и мелиорации. Т. III, 1927.
Берг Л. С. О виде и его подразделениях. (А. Семенов-Тянь-Шанский Таксономические границы вида и его подразделений. Опыт точной категорп зации низших систематических единиц). Записки АН, физико-математический отдел. Т. 25, № 1, 1910.
Берг Л. С. Фауна России. Изд. Академии Наук. Т. I. СПб, 1911.
Берг Л. С. Каспийские сельди, собранные Каспийской научно-промысловой экспедицией 1912 г. вдоль западного берега моря. Материалы к познанию русского рыболовства. Т. II. Вып. 3, 1913.
Берг Л. С. О распространении речного угря (Anguilla anguilla L.) и России. Ежегодник Зоологического музея Академии Наук. Т. 21. 1916.
Берг Л. С. Очерк промысловых исследований в России. Известия Государственного института опытной агрономии. Т. II, Вып. 3, 1924.
Берг Л. С. Яровые и озимые расы у проходных рыб. Известия АН СССР. Отдел математики и естественных наук, 1934.
Берг Л. С. Яровые и озимые расы у рыб. «Природа», 1934, № 4.
Берг Л. С. Успехи ихтиологии 1917-1937 гг. Известия АН СССР, серии биологии,№ 5, 1937.
Берг Л. С. Рыбы. В кн. «Жизнь пресных вод СССР». Т. I, Изд. AН СССР, 1940.
Берг Л. С. Система рыбообразных и рыб, ныне живущих и ископаемых. Труды Зоологического института АН СССР. Т. 5. Вып. 2, 1940.
То же. Изд. 2-е. Там же. Т. 20, 1955.
Берг Л. С. Рыбы Финского залива. Известия ВНИОРХ. Т. 23, 1940.
Берг Л. С. Рыбы пресных вод СССР и сопредельных стран. Изд. АН СССР. Ч. I-III, 1948-1949.
Берг Л. С. и Световидов А. Н. Рыбы. В кн. «Успехи биологических наук в СССР за 25 лет (1917-1942)». Изд. АН СССР, 1945.
Берг Л. С. и Правдин И. Ф. Рыбы Кольского полуострова. Известия ВНИОРХ. Т. 26. Вып. 2, 1948.
Бердичевский Л. С. Методика составления промысловых карт. Труды Совещаний. Вып. 5, Изд-во АН СССР, 1955.
Богоров В. Г. Исследования питания планктоноядных рыб. Бюллетень ВНИРО, № 1, 1934.
Бойко Е. Г. Методика определения возраста рыб по спилам плавников. Труды АзчерНИРО. Т. 5, 1951.
Бокова Е. Н. Методика изучения питания рыб в естественных условия на ранних этапах развития. Труды Совещания по методике изучения кормовой базы и питания рыб. М., Изд. АН СССР, 1955.
Болховитянов В. Д. Как составить промысловую карту и определить размер сырьевых ресурсов рыбного хозяйства Дальнего Востока, «За социалистическое рыбное хозяйство», № 8-9, «Социалистическая реконструкции рыбного хозяйства Дальнего Востока», 1931, № 11-12.
Борзенко М.П. Материалы по биологии сазана. Известия Бакинской ихтиологической лаборатории. Вып. 2 (1), 1926.
Борзенко М. П. Материалы по биологии сазана. Известия Бакинской ихтиологической лаборатории. Т. 2, Вып. 1, 1935.
Борисов П. Г. Рыбы реки Лены. Труды Комиссии по изучению Якутской АССР. Т. 9, 1928.
Борисов П. Г. Научно-промысловые исследования на морских и пресных водоемах. Изд. 2-е. М., «Пищевая промышленность», 1964.
Борисов П. Г. и Богданов А. С. Сырьевая база рыбной промышленности СССР. М., Пищепромиздат, 1955.
Борисов П. Г. и Овсянников Н. С. Определитель промысловых рыб. М., Пищепромиздат, 1958.
Борисов П. Г. и Никольский Г. В. Основные этапы развития исследований отечественных биологических рыбохозяйственных 100 лет. Зоологический журнал. Т. 40. Вып. 8, 1961.
Бородин Н. А. Исследование образа жизни каспийских сельдей. «Вестник рыбопромышленности», 1904, № 7.
Боруцкий Е. В. К вопросу о годовых колебаниях весенней биомассы бентоса в озерах. Зоологический журнал. Т. 25. Вып. 5, 1946.
Боруцкий Е. В. Методика изучения питания растительноядных рыб. Труды Совещания по методике изучения кормовой базы и питания рыб. М., Изд. АН СССР, 1955.
Браунер А. Заметки о сельдях Черного и Азовского морей. Труды Бессарабского общества естествоиспытателей. Т. II. Вып. 2, 1912.
Броцкая В. А Инструкция для сборов и обработки материалов по питанию бентосоядных рыб. М., Пищепромиздат, 1939.
Букирев А. И. К географической изменчивости пеляди – Coregonus peled (Gmelin). Известия биологического института при Пермском государственном университете. Т. XI. № 7-8, 1938.
Булгаков Г. П. К познанию каспийских сельдей. Описание нового вида рода Caspialosa. Бюллетень Среднеазиатского госуниверситета № 13 1926.
Булгаков Г. П. К систематике каспийских сельдей. Бюллетень САГУ, № 16, 1927.
Бурмакин Е.В. Состав ихтиофауны бассейна Гыданского залива. «Проблемы Арктики», 1938, № 3.
Бурмакин Е.В. Биология и рыбохозяйственное значение пеляди. Труды Барабинского отделения ВНИОРХ. Т. 6. Вып. I. Новосибирск, 1953.
Бурмакин Е.В. Результаты работы по обезрыбливанию двух озер Ленинградской области. Научно-технический бюллетень ГосНИОРХ, № 12, 1960.
Бурмакин Е.В. Предварительные результаты выращивания пеляди и карпа в озере, обезрыбленном полихлорпиненом. Научно-технический бюллетень ГосНИОРХ, № 15, 1962.
Бурмакин Е.В. Химический метод рыбохозяйственного преобразования озер (Предварительные результаты и перспективы). Известия ГосНИОРХ. Т. 55, 1963.
Бурмакин Е. В. Акклиматизация пресноводных рыб в СССР. Известия ГосНИОРХ. Т. 53, 1963.
Бурмакин Е. В. и Домбровский Г.В. Состояние рыбных запасов озера Балхаша и перспективы увеличения уловов. Известия ВНИОРХ. Т. 37, 1956.
Бурмакин Е.В. и Тюрин П. В. О биологической классификации рыб. «Вопросы ихтиологии». Вып. 13, 1959.
Бурмакин Е.В. и Жаков Л. А. Опыт определения рыбопродуктивности окуневого озера. Научно-технический бюллетень, ГосНИОРХ» № 13-14. 1961.
Вальмус С. Г. и Правдин И.Ф. Материалы по обследованию Галичского озера. Труды Галичского отделения Костромского научного общества no изучению местного края. Вып. 2, 1923.
Варпаховский Н А. Определитель пресноводных рыб Европейской России. СПб, 1898.
Варпаховский Н. Рыболовство в бассейне Оби. И. Рыбы бассейна р, Оби. СПб. 1902.
Васнецов В. В. Дивергенция и адаптация в онтогенезе. Зоологический журнал. Т. 25. Вып. 3, 1946.
Васнецов В. В. Рост как адаптация. Бюллетень Московского общество испытателей природы. Т. III. Вып. I, 1947.
Васнецов В. В. (ред.) Морфологические особенности, определяющие питание леща, воблы и сазана на всех стадиях развития. Изд. АН СССР.
Васнецов В. В. Опыт анализа роста рыб реки Амура. Труды Амурской ихтиологической экспедиции. Т. 4, 1951.
Васнецов В. В. О закономерностях роста рыб. Очерки по общим вопросам ихтиологии. Изд. АН СССР, М.-Л., 1953 а.
Васнецов В. В. О морфологии. Зоологический журнал. Т. 32. Вып. 6, 1953 б.
Васнецов В. В. Этапы развития костистых рыб. Очерки по общим вопросам ихтиологии. Изд. АН СССР, 1953 в.
Васнецов В. В. Происхождение нерестовых миграций проходных рыб. Очерки по общим вопросам ихтиологии. Изд. АН СССР, 1953 г.
Веденскии А. П. Опыт поисков минтая по плавающей икре Труды ТИНРО. Т. 24, 1949.
Великохатько Ф. Д. Миграционные пути проходных рыб Черного моря и их исторические причины. Зоологический журнал. Т. 17, Вып. 5, 1938.
Великохатько Ф.Д. Материалы к познанию леща из р. Днепра. Зоологический журнал. Т. 20. Вып. 1, 1941.
Веригин Б. В. Возрастные изменения молоди толстолобика в связи с его биологией. Труды Амурской ихтиологической экспедиции 1945-1949 гг., 1950.
Верещагин Г. Ю., Аничков Н.А., Предтеченский Н.П. и др. Опыт гидрологической съемки озера Ласси-Лампи Ленинградского уезда, Токсовской волости. Сборник в честь Н. М. Книповича, 1927.
Вернидуб М. Ф. Материалы по систематике, биологии и промыслу дальневосточных палтусов. Изд. Ленинградского государственного университета, 1935.
Вернидуб М. Ф. Материалы к познанию тихоокеанского палтуса. Труды Ленинградского общества естествоиспытателей. Т. 15. Вып. 2, 1936.
Вернидуб М.Ф.О морфофизиологических этапах в развитии личинок рыб. Доклады АН СССР. Т. 21, № 3, 1950.
Вильсон А. П. Мечение кандалакшских и атлантическо-скандинавских сельдей, «Рыбное хозяйство», 1956, № 7.
Виноградова 3. А. О явлении линьки у некоторых рыб Черного моря. Труды Карадагской биологической станции. Вып. 9, 1950.
Виролайнен М.П. Нерестилища семги на р. Кеми. Труды Карело-Финского отделения ВНИОРХ. Т. II, 1946.
Владимиров В. И. Ручьевая форель Армении и ее отношение к другим представителям рода Salmo. Труды Севанской гидробиологической станции. Т. 10, 1948.
Владимиров В. И. Условия размножения рыб в нижнем Днепре и Каховском водохранилище. Киев. Изд. АН УССР, 1955.
Владимиров В. И., Сухойван П. Г., Бугай К. С. Размножение рыб в условиях зарегулированного стока реки (на примере Днепра). Киев. Изд. АН УССР, 1963.
Владимирская М. И. Нерестилища семги в верховьях р. Печоры и меры для увеличения их производительности. Труды Печоро-Илычского государственного заповедника. Сыктывкар, 1957.
Вовк Ф. И. О методике реконструкции роста рыб по чешуе. Труды биологической станции Борок АН СССР. Т. 2, 1955.
Вукотич Н.Н. К определению плодовитости сельдей из рода Caspialosa. Материалы к познанию русского рыболовства. Т. IV. Вып. 6, 1915.
Галкин Г. Г. Атлас чешуи пресноводных костистых рыб. Известия ВНИОРХ. Т. 46, 1958.
Гербильский Н. Л. Биологические группы куринского осетра (Acipenser guldenstadti persicus Borodin) и основания для их заводского воспроизводства. ДАН СССР. Т. 71, № 4, 1950.
Гербильский Н. Л. Внутривидовые биологические группы осетровых и их воспроизводство в низовьях рек с зарегулированным стоком. «Рыбное хозяйство». 1953, № 4.
Гершензон С.М., Геронимус Е.С. и Аврех В. В. Серологический анализ генотипов дрозофилы. Журнал микробиологии, эпидемиологии и иммунологии. Т. 16. Вып. 5, 1936, с. 763-770.
Гинзбург Я. И. Развитие сома. Труды Каспийского бассейнового” филиала ВНИРО. Т. II, 1950.
Голованов Ф. Северо-каспийский осетр. «Рыбное хозяйство», 1940, № 5
Головков Г. А. и Кузьмин А. Н. Биология пеляди и биотехника ее разведения. Изд-во «Рыбное хозяйство», 1963.
Гомоюнов К. А. Гидрологический режим бухты Патрокл в связи с метеорологическими условиями. Известия Тихоокеанской научно-промысловой станции. Т. 1. Вып. 2, 1928.
Гончаров Г. Д. Серологическая диагностика, как новое доказательство вирусной природы краснухи карпов. «Рыбное хозяйство», 1949, № 4.
Гончаров Г. Д. К диагностике краснухи карповых рыб. «Рыбное хозяйство», 1950, № 9.
Гончаров Г. Д. Реакция агглютинации при краснухе карповых рыб. Труды ВНИРО. Т. 19, 1951 а.
Гончаров Г. Д. К иммунизации рыб. Доклады АН СССР, Новая серия. Т. 78, № 3, 1951 б.
Гончаров Г. Д. Вирусная краснуха рыб в СССР и за рубежом. Труды совещания по болезням рыб. В кн. «Совещание по болезням рыб», 1959.
Гончаров Г. Д. Иммунологическая реактивность у рыб. Бюллетень института биологии водохранилищ. АН СССР, № 12, 1962.
Горский Н. В. Семужий промысел на реке Выг. В кн. «Рыбное хозяйство Карелии». Вып. 1, 1932.
Горский Н. В. Семга реки Выг, ее промысел и биология. Труды Карельской научно-исследовательской рыбохозяйственной станции. Т. I, 1935.
Гриб А. В. Некоторые особенности в постэмбриональном развитии вьюна (Misgurnus fossilis L., сем. Cobitidae, Cyprinoidea). Труды Ленинградского общества естествоиспытателей. Т. 65. Вып. 2, 1936.
Гримм О. А. . О происхождении тупо- и острорылых разновидностей стерляди, «Вестник рыбопромышленника». 1892, № 1.
Гудимович П. К. Краткие сведения о плодовитости черноморской султанки и характер ее икрометания. Зоологический журнал. Т. 30. Вып. I, 1952.
Дементьева Т. Ф. и Ильин Б. С. Инструкция по сбору и первичной обработке ихтиологических материалов. Пищепромиздат, 1938.
Дементьева Т.Ф. Распределение и миграция воблы в море. Труды ВНИРО. Т. 10. Ч. I, 1939.
Дементьева Т.Ф. Методика составления прогноза уловов леща Северного Каспия. Труды ВНИРО. Т. 21, 1952.
Державин А. Н. Севрюга (Acipenser stellatus). Биологический очерк. Известия Бакинской ихтиологической лаборатории. Т. 1, 1922.
Державин А.Н. Рыбные запасы и рыбохозяйственная мелиорация Каспия. Бюллетень Всекаспийской экспедиции, № 5-6, 1932.
Державин А. Н. Воспроизводство запасов осетровых рыб. Баку. Изд. АН Азербайдж. ССР, 1947.
Дерюгин К.М. Новая форма трески из Могильного озера. Труды Петроградского общества естествоиспытателей. Т. 51. Вып. 1, 1920.
Дерюгин К.М. Биология материковых вод. Исследование бентоса. Изд. Государственного гидрологического института, 1931.
Детлаф Т. А. и Гинзбург А. С. Зародышевое развитие осетровых рыб (севрюги, осетра и белуги) в связи с вопросами их разведения. Изд. АН СССР, 1954.
Дехтярева А. И. Опознавательные признаки сеголетков сельдей Северного Каспия. Труды. ВНИРО. Т. 14, 1940.
Джонстоя Д. Условия жизни в море (перевод Книповича Н. М.), Пг, 1919.
Диксон Б. И. Результаты наблюдений над биологией черноспинки в 1905 г. «Вестник рыбопромышленника», 1905, № 11.
Догель В. А. Общая характеристика паразитофауны рыб северной Карелии. Труды 1-й научно-технической конференции по рыбной промышленности Карело-финской ССР, 1947.
Дойников К. Г. Материалы по биологии и оценке запасов осетровых рыб Азовского моря. Работы Доно-Кубанской научно-рыбохозяйственной станции. Вып. 4, 1936.
Домрачев П. Ф. К вопросу о классификации озер Северо-западного края. Известия Государственного гидрологического института № 2, 1923.
Домрачев П. Ф. Схема измерения сигов и результаты биометрических измерений волховского сига. Материалы по исследованию Волхова. Вып. 3. Ч. 1, 1924.
Дорошин Г. Я. и Троицкий С. К. Характеристика условий размножения кубанской севрюги в 1944-1947 гг. Труды рыбоводно-биологической лаборатории Азчеррыбвода. Вып. 1, 1949.
Дружинин А. Д. Миграция сельди в водах Сахалина по данным опытов мечения 1956-1958 гг. «Рыбное хозяйство», 1959, № 9.
Дрягин П. А. Рыбные ресурсы Якутии. Труды СОПС АН СССР. Якутская АССР. Вып. 5, 1933.
Дрягин П. А. Размеры рыб при наступлении половозрелости. «Рыбное хозяйство», 1934, № 4.
Дрягин П. А. Рыбы реки Чу и рыбное хозяйство этой реки. Труды Киргизской комплексной экспедиции 1932-1933 гг. Т. 3, Вып. 1, 1936.
Дрягин П. А. Основной метод исследования нерестилищ озерных рыб. «Рыбное хозяйство», 1938, № 12.
Дрягин П. А. Нерест рыб на оз. Ильмене. Информационный бюллетень ВНИОРХ, № 3, 1939.
Дрягин П. А. Порционное икрометание у карповых рыб. Известия ВНИОРХ. Т. 21, 1939.
Дрягин П. А. О яловости у рыб. Информационный сборник консультационного бюро ВНИОРХ, № 4, 1940.
Дрягин П. А. Об определении потенционального роста и потенциональных размеров рыб. Известия ВНИОРХ. Т. 26, Вып. 1, 1947.
Дрягин П. А. Промысловые рыбы Обь-Иртышского бассейна. Известия ВНИОРХ. Т. 25, Вып. 2, 1948.
Дрягин П. А. Основания к регулированию роста рыб. Бюллетень рыбного хозяйства Карело-Финской ССР, № 3, 1948.
Дрягин П. А. Биоэкологические группы рыб и их происхождение. Доклады АН СССР. Т. 66, № 1, 1949.
Дрягин П. А. Половые циклы и нерест рыб. Известия ВНИОРХ. Т. 28, 1949.
Дрягин П. А. О методах учета рыбопромысловых запасов в пресноводных водоемах. Труды проблемных и тематических совещаний. Вып. 1. Проблемы гидробиологии внутренних вод. 1. Изд. АН СССР, 1951.
Дрягин П. А. О полевых исследованиях размножения рыб. Известия ВНИОРХ. Т. 30, 1952.
Дрягин П. А Акклиматизация рыб во внутренних водоемах СССР. Известия ВНИОРХ. Т. 32, 1953.
Дрягин П. А. Биологические основы реконструкции фауны рыб в озерах СССР. Пищепромиздат, 1956.
Дрягин П. А. Основные направления в изучении жизненных циклов рыб. Научно-технический бюллетень ГосНИОРХ, № 13-14, 1961.
Дрягин П. А. Состояние и основные задачи ихтиологических исследований в СССР. Вторая зоологическая конференция БССР, Минск, 1962.
Европейцева Н.В. Расовый анализ беломорской трески. Изд. Ленинградского университета, 1937.
Европейцева Н. В. Возрастные адаптации в жаберном аппарате Percidae в связи с возрастными изменениями характера питания. Ученые записки Ленинградского государственного университета, № 35, 1939.
Европейцева Н. В. Личиночный период налима. Труды Ленинградского общества естествоиспытателей. Т. 69. Вып. 5, 1946.
Европейцева Н.В. Морфологические черты постэмбрионального развития сигов. Труды Лаборатории основ рыбоводства. Т. 2, 1949 а.
Европейцева Н.В. Морфологические черты постэмбрионального развития окуневых. Ученые записки Ленинградского госуниверситета. Серия
биологических наук. Вып. 23, 1949 б.
Егоров А. Г. Байкальский осетр. Систематика, биология, промысел, сырьевая база и воспроизводство запасов. Улан-Удэ, АН СССР, 1961.
Елеонский А.Н. Прудовое рыбоводство. М., 1946.
Есипов В. К. О треске Баренцева моря. «Карело-Мурманский край», Мурманск, 1931, № 9-10.
Есипов В. К. Указатель литературы по рыбному хозяйству Европейского Севера СССР. 1917-1933 гг. Л. Изд. Главсевморпути. 1933.
Есипов В. К. Указатель литературы по рыбному хозяйству Европейского Севера СССР 1917-1933 гг. Л., 1935.
Есипов В. К. Рыбы Карского моря. Изд. АН СССР, 1952.
Ефимова А. И. Щука Обь-Иртышского бассейна. Известия ВНИОРХ.Т. 28, 1949.
Ефимова А. И. Возраст наступления половой зрелости у костистых рыб – важнейший показатель численности их. Известия ВНИОРХ. Т. 32,1954.
Жадин В. И. Методика изучения донной фауны водоемов и экологии
водных беспозвоночных. В кн. «Жизнь пресных вод СССР», Т. 4. Ч. 1, 1956.
Жадин В. И. Методы гидробиологического исследования. «Высшая школа»,1960.
Жадин В. И. и Герд С. В. Реки, озера и водохранилища СССР, ихфауна и флора. М., Учпедгиз, 1961.
Жаков Л. А. Способ определения абсолютной численности рыб посредством подсчета окуневых кладок. Озера Карельского перешейка. Изд. АН
СССР, М.-Л., 1964.
Желтенкова М.В. Питание воблы в северной части Каспийского моря. Труды ВНИРО. Т. 10. Ч. 1, 1939.
Жуков П. И. Рыбы бассейна Немана (в пределах Белорусской ССР). Изд. АН БССР, 1958.
Жуков П. И. Определитель рыб БССР. (Справочник для практических работников). Минск, изд. АН БССР, 1960.
Зайцев В. П. и Радаков Д. В. Использование подводной лодки для научных рыбохозяйственных исследований. Сб. «Советские рыбохозяйственные исследования в морях Европейского Севера». М., 1960.
Закс И. Г. Предварительные данные о распределении флоры и фауны в прибрежной полосе залива Петра Великого в Японском море. Производительные силы Дальнего Востока. Т. V. Животный мир, 1927.
Закс И. Г. К познанию донных сообществ Шантарского моря. Известия Тихоокеанской научно-промысловой станции. Т. 3. Вып. 2, 1929.
Закс М. Г. и Соколова М. М. Установление различий между отдельными стадами нерки (Oncorhynchus nerka Wabb.) посредством реакции преципитации. Доклады АН СССР. Т. 139, № 6, 1961.
Замахаев Д.Ф. Нерестовые марки на чешуе каспийских сельдей. Труды ВНИРО. Т. 14, 1940.
Замахаев Д.Ф. К методике расчисления роста трески по отолитам. Зоологический журнал. Т. 20. Вып. 2, 1941.
Замахаев Д.Ф. К вопросу о систематическом положении проходных сельдей Каспия. Зоологический журнал. Т. 23. Вып. 2-3, 1944.
Замбриборщ Ф.С. О некоторых анатомических признаках черноморских кефалей. Зоологический журнал. Т. 30. Вып. 2, 1951.
3инова Е. С. Морская капуста (Laminaria) и др. водоросли, имеющие промышленное значение. Известия Тихоокеанской научно-промысловой станции. Т. 2. Ч. 2, 1928.
3ограф Н. Ю. Материалы к познанию организации стерляди. Известия общества любителей естествознания, антропологии и этнографии. Т. 13. Вып. 3, 1887.
3убович П. О. К вопросу о черноморских бычках. Труды Всеукраинской государственной Черноморско-Азовской научно-промысловой станции. Вып. 1, 1925.
Иванова-Берг М.М. Наблюдения над весенним ходом и нерестом невской миноги. Известия АН СССР, № 2-3, 1936.
Ивлев В. С. Метод оценки популяционной плодовитости рыб. Труды Латвийского отделения ВНИРО. Т. 1, 1953.
Ильин Б. С. Определитель бычков (fam. Gobiidae) Азовского и Черного морей. Труды Азовско-Черноморской научно-промысловой экспедиции. Вып. 2, 1927.
Ильин Б. С. и Певзнер В. И. Новые сведения о беломорских видах трески. Сборник в честь Н. М. Книповича. М., Пищепромиздат, 1939.
Иоганзен Б. Г. Гидробиологические исследования водоемов в окрестностях Новосибирска. Бюллетень Зоологической секции Томского общества испытателей природы, № 1, 1937.
Иоганзен Б. Г. Вопросы теории и практики интродукции рыб. Труды Томского госуниверситета. Т. 97, 1946.
Иоганзен Б. Г. Рыбы бассейна реки Оби. Изд. Томского государственного университета, 1948.
Иоганзен Б. Г. Опыт изучения численности проходных сиговых и интенсивности рыболовства средней Оби. Труды Томского государственного университета. Т. 119, 1952.
Иоганзен Б. Г. К изучению плодовитости рыб. Труды Томского государственного университета. Т. 121, 1955.
Иоганзен Б. Г. и ПеткевичА. Н. Акклиматизация рыб в Западной Сибири. Труды Барабинского отделения ВНИОРХ. Т. 5, 1951.
Исаченко В. А. Новый вид сига из бассейна р. Енисея. Труды Сибирской ихтиологической лаборатории. Т. 2, Вып. 2, 1925.
Кагановская С.М. Метод определения возраста и состав уловов колючей акулы. Вестник Дальневосточного филиала АН СССР, № 1-3, 1933.
Кагановская С. М. Материалы к промысловой биологии колючей акулы. Известия ТИНРО. Т. 10, 1937.
Кагановский А. Г. Некоторые вопросы биологии и динамики численности горбуши. Известия ТИНРО. Т. 31, 1949.
Кагановский А. Г. Миграция скумбрии в Японском море. Известия ТИНРО. Т. 35, 1951.
Казанский В. И. Материалы по развитию и систематике личинок карповых рыб. Труды ихтиологической лаборатории. Т. 3. Вып. 7. Астрахань, 1915.
Казанский В. И. Схема определения молоди карповых рыб. Бюллетень рыбного хозяйства, № 6-7, 1924.
Казанский В. И. Этюды по морфологии и биологии личинок рыб Нижней Волги. Труды ихтиологической лаборатории, Т. 5. Вып. 3, Астрахань,, 1925.
Казанова И. И. и Халдинова Н. А. Места и условия нерест каспийских сельдей в дельте Волги (по распределению их икры и личинок). Труды ВНИРО, Т. 14, 1940.
Казанчеев Е. Н. Рыбы Каспийского моря. М., «Рыбное хозяйство», 1963.
Караваев Г. А. Миграции воблы Rutilus rutilus caspicus Jak. в Северном Каспии. Труды ВНИРО. Т. 10. Ч. 1, 1939.
Караваев Г. А. Инструкция по мечению рыб. Изд. ВНИРО, 1958.
Карпевич А. Ф. Теоретические предпосылки к акклиматизации водных организмов. Труды ВНИРО. Т. 43. Вып. 1, 1960.
Карантонис Ф. Э. Морфологическая характеристика популяций сельди (Clupea harengus harengus L.) Баренцева моря. Труды ПИНРО. Т. 1, 1938
Карантонис Ф. Э. Новое в изучении чешуи сазана. Научный бюллетень Ленинградского университета, № 3, 1945.
Карзинкин Г. С. Использование радиоактивных изотопов в рыбном хозяйстве. М., Пищепромиздат, 1962.
Карзинкин Г. С. Значение химикатов в рыбном хозяйстве. «Вопросы ихтиологии». Т. 4. Вып. 2 (3), 1964.
Кесслер К. Описание рыб, которые встречаются в водах С.-Петербургской губернии. Изд. Русского энтомологического общества, 1864.
Кесслер К. Описание рыб, принадлежащих к семействам, общим Черному и Каспийскому морям. Труды С.-Пб. общества естествоиспытаний. V, 1874.
Кесслер К. Ф. Рыбы, водящиеся и встречающиеся в Арало-Каспийско- Понтийской ихтиологической области. Труды Арало-Каспийской экспедиции. Т. 4, С.-Пб., 1877.
Кирпичников В. С. Световидов А. Н. и ТрошинА. С. Мечение карпа радиоактивными изотопами фосфора и кальция. ДАН СССР. Т. 111, № 1, 1956.
Кирпичников В. С., Световидов А. Н., Трошин А. С. Мечение рыб радиоактивными изотопами фосфора и кальция. Труды Совещания по физиологии рыб. (1956) М., Изд. АН СССР, 1958.
Киселев О. Н. и Соловьев Б. С. Результаты наблюдений за поведением рыб из глубоководного гидростата. «Вопросы ихтиологии». Т. 1. Вып. 4 (2), 1961.
Киселевич К. А. Каспийско-волжские сельди. 1. Систематика. Труды Астраханской научно-промысловой экспедиции 1914-1915 гг., 1923а.
Киселевич К. А. Материалы по биологии каспийских сельдей. II. О плодовитости каспийско-волжских сельдей. Труды Астраханской ихтиологической лаборатории. Вып. 1, 19236.
Киселевич К. А. Исследовательская работа в Волго-Каспийском районе (Ихтиологическая лаборатория, ее возникновение, развитие и достижения). «Наш край». Астрахань, 1927, № 8.
Киселевич К. А. Сельди северного Каспия. Обл. изд-во, Сталинград, 1937.
Кленова М. В. Отчет о работе комиссии по механическому анализу грунтов при Государственном океанографическом институте. Бюллетень Госуд. океанографического института. Вып. I, 1931.
Клер В. О. Некоторые данные к определению возраста рыб по костям. «Вестник рыбопромышленности», 1916, № 3.
Клер В. О. Метод шлифов в применении микроструктуры древесного угля, костей и древесины. Известия Уральского политехнического института, Т. 6,1927а.
Клер В. О. К методике исследования периодики роста. Метод изодинамических плоскостей. Зоологический журнал. Т. 7. Вып. 4, 19276.
Книпович Н. М. Экспедиция для научно-промысловых исследований у берегов Мурмана. Т. 1, Спб, 1902.
Книпович Н. М. Гидрологические исследования в Каспийском море в 1914-1915 гг. Труды Каспийской экспедиции 1914/1915, 1921.
Книпович Н. М. Определитель рыб Черного и Азовского морей. М., 1923.
Книпович Н. М. Определитель рыб морей Баренцева, Белого и Карского. Труды института по изучению Севера. Вып. 27, 1926.
Книпович Н. М. Гидрология морей и солоноватых вод. М.-Л., Пищепромиздат, 1938.
Книпович Н. М., Арнольд И. Н. и Бородин Н. А. От комиссии по собиранию и изучению рыбьей молоди при Российском обществе во рыбоводству. «Вестник рыбопромышленности», 1904, № 7.
Коблицкая А. Ф. Изучение нерестилищ пресноводных рыб (методическое пособие). Астрахань, Изд. «Волга» (Астраханский государственный заповедник), 1963.
Кожина Е. С. Наблюдения над ранними стадиями жизни леща в Миккельском озере и Крошнозере. Материалы по повышению рыбной продуктивности малых озер Карелии. Труды Карельского филиала АН СССР Вып. II, 1956.
Кожов М. М. и Мишарин К. И. (ред). Рыбы и рыбное хозяйство в бассейне озера Байкал. 1958.
Константинов К. Г. Мечение промысловых рыб. Мурманск, 1956. Константинов К. Г. Результаты мечения донных рыб Баренцева моря в 1946-1955 гг. Труды ПИНРО. Вып. 10, 1957.
Кохиеико С. В. Угорь в водоемах Белорусской ССР. Известия АН БССР, 6, 1954.
Кохиеико С. В. Биология и распространение угря. Минск, Изд. АН Белорусской ССР, 1958.
Кохненко С. В. и Боровик Е. А. Морфологическая характеристика стекловидных угорьков. Минск, Изд. АН Белорусской ССР, 1957.
Кошелев Б. В. Изменения половых циклов у рыб с единовременным икрометанием в связи с изменением условий существования. «Вопросы ихтиологии». Т. 1. Вып. 4 (21), 1961.
Крашенинников С. Описание Земли Камчатки. Т. 1. Изд. АН СПб, 1755.
Крохин Е. М. и Крогиус Ф. В. Очерк бассейна реки Большой и нерестилищ лососевых. Известия ТИНРО. Т. 9., 1937.
Крыжановский С. Г. Экологические группы рыб и закономерности их развития. Известия ТИНРО. Т. 27, 1948.
Крыжановский С. Г. Эколого-морфологические закономерности развития карповых, вьюновых и сомовых рыб. Труды Института морфологии животных. Вып. 1, 1949.
Крыжановский С. Г. Особенности зрелых яиц костистых рыб. «Вопросы ихтиологии». Вып. 1, 1953.
Крыжановский С. Г. Материалы по развитию сельдевых рыб. Труды Института морфологии животных. Вып. 17, 1956.
Крыжановский С. Г., Смирнов А. И., Соин С. Г. Материалы по развитию рыб Амура. Труды Амурской ихтиологической экспедиции 1945-1949 гг. Т. 2, 1951.
Крыжановский С. Г., Дислер Н. Н., Смирнова Е. Н. Эколого-морфологические закономерности развития окуневидных рыб. Труды Института морфологии животных. Вып. 10, 1953.
Крыжановский С. Г. и Троицкий С. К. Материал об ихтиофауне рек Черноморского побережья. (В пределах Краснодарского края). «Вопросы ихтиологии». Вып. 2, 1954.
Кудерский Л. А. Материалы по внутривидовой изменчивости судака. Труды Карельского филиала АН СССР. Вып. 13, 1958.
Кудерский Л. А. Условия существования и перспективы расселения судака водоемов Карелии. «Рыбное хозяйство Карелии». Вып. 8, 1964.
Кузнецов И. И. Материалы к искусственному разведению кеты на Амуре в 1909-1910 гг. Материалы к познанию русского рыболовства Т. 50. Вып. 3, 1912.
Кулаев С. И. Наблюдения над изменением семенников речного окуня в течение годового цикла. Русский зоологический журнал. Т. VII. Вып. 3, 1927.
Кулаев С. И. Годовой цикл и шкала зрелости половозрелой плотвы {Rutilus rutilus L.). Записки Болшевской биологической станции. Вып. 2, 1939.
Куликова Е Б. Рост и возраст глубоководных рыб. Труды Института океанологии АН СССР. Т. 20, 1957.
Лагунов И. И. Молодь лососей рода Oncorhynchus. Зоологический журнал. Т. 18. Вып. 5, 1939.
Лапин Ю. Е. Типы нерестовых популяций и некоторые вопросы методики исследования динамики численности рыб. «Вопросы ихтиологии». Т. 1. Вып. 4 (21), 1961.
Ларина Н. И. и Кулькова А. В. К вопросу о кровном родстве близких видов и географически удаленных популяций. Совещание по экологической физиологии. Тезисы докладов. Т. 1. Изд. АН СССР, 1959.
Ламперт. Жизнь пресных вод. СПб, 1900.
Лапицкий И. И. Овогенез и годичный цикл яичников у сига лудоги. Труды лаборатории основ рыбоводства. Т. II, 1949.
Ласточкин Д. А. Стоячие водоемы (озера и пруды). Руководство для школьных экскурсий и краеведных исследований. Иваново-Вознесенск. Изд-во «Основа», 1925.
Лебедев Н. В. Элементарные популяции рыб. Зоологический журнал. Т. 25, Вып. 2, 1946.
Леванидов В. Я. Питание тайменя в предгорных притоках Амура. Бюллетень Московского общества испытателей природы. Отдел биологии. Т. 56. Вып. 6, 1951.
Лепнева С. Г. Жизнь в озерах. Сб. «Жизнь пресных вод СССР». Т. III, 1950.
Летичевский М. А. К вопросу о плодовитости рыб юга Аральского моря. Зоологический журнал. Т. 25. Вып. 4, 1946.
Линдберг Г. У. Список рыб, собранных А. А. Емельяновым в реках Ботчи и Копи, впадающих в Японское море, с описанием нового вида из рода Coitus (С. emeljanovi). Труды Дальневосточного университета, серия VIII, №2, 1927.
Линдберг Г. У. Успехи ихтиологии за 20 лет. «Природа», 1937, № 10.
Линдберг Г. У. О систематике и распространении песчанок рода Ammodytes. Вестник Дальневосточного филиала АН СССР, № 27, 1937.
Линдберг Г. У. О родах и видах рыб семейства Blenniidae близких К роду Anoplarchus. Труды Гидробиологической экспедиции Зоологического музея АН СССР в 1934 г. на Японском море. Вып. 1, Дальневосточный филиал АН СССР, 1938.
Линдберг Г. У. Новый принцип перевозки живой рыбы. «Рыбное хозяйство». 1946, № 7.
Линдберг Г. У. Основные принципы и методы составления рыбопоисковых карт для дальневосточных морей. Труды проблемных и тематических совещаний. Вып. 6. Изд. АН СССР, М., 1956.
Линдберг Г. У. и Охрямкин Д. И. О камбалах Японского моря. Доклады АН СССР. Т. 4/13, № 3, 1936.
Липина Н. Н. Личинки и куколки хирономид. Экология и систематика, 1929.
Лишев М. Н. К методике изучения состава пищи хищных рыб. Известия ТИНРО. Т. 32, 1950.
Лужин Б. П. Иссык-кульская форель гегаркуни. Фрунзе, 1956.
Лукаш Б. С. Рыбы реки Вычегды. «Север». Вятка, 1923.
Лукаш Б. С. Материалы к филогении карповых рыб (сем. Cuprinidae). Труды Карельского филиала Академии наук СССР. Вып. 5, 1956.
Лукаш Б.С. Рыбы верховьев р. Камы. Материалы Верхнекамской ихтиологической экспедиции, 1929.
Лукаш Б. С. Рыбы нижнего течения реки Вятки. Труды Вятского научно-исследовательского института краеведения. Т. 6, 1938.
Лукин А. В. К биологии волжского налима. Труды Татарского отделения ВНИОРХ. Т. 2, 1935.
Лукин А. В. О роли температурных факторов в процессе приспособления размножения рыб к условиям среды. Зоологический журнал. Т. 58, № 4, 1947.
Лукин А. В. Зависимость плодовитости рыб и характера их икрометания от условий обитания. Известия АН СССР, серия биологии, № 5, 1948.
Лукин А. В. Основные черты экологии осетровых в Средней Волге. Труды Татарского отделения ВНИОРХ. Вып. 5, 1949.
Лукьяненко В. И. и Миэсерова Е. К. Сравнительно-иммунологическое изучение комплементарной функции крови рыб. Доклады АН СССР. Т. 146, № 4, 1962.
Майорова А. А. Таксономическое положение хамсы, ловимой у берегов Грузии. Труды рыбохозяйственной и биологической станции Грузии. Т. 1. Вып. 1, 1934.
Майр Э. Систематика и происхождение видов. Пер. с анг. М., 1947.
Майр Э., Линсли Э. и Юзингер Р. Методы и принципы зоологической систематики. Пер. с англ., М., 1956.
Майский В. Г. К методике определения рыбных запасов в Азовском море. «Рыбное хозяйство», № 3, 1939.
Мантейфель Б. П. Краткая характеристика основных закономерностей в изменениях планктона Баренцева моря. Труды ПИНРО. Т. 1, 1938.
Мантейфель Б. П. Адаптивное значение периодических миграций водных организмов. «Вопросы ихтиологии». Вып. 13, 1959.
Мантейфель Б. П. и Марти Ю. Ю. Исследование нереста мурманской сельди. Труды ПИНРО. Т. 4, 1939.
Маркун М. И. К систематике и биологии налима р. Камы. Известия Пермского биологического научно-исследовательского института. Т. 10. Вып. 6, 1936.
Маслов Н. А. Промысловые донные рыбы Баренцева моря. Труды ПИНРО. Вып. 8, 1944.
Марти Ю. Ю. Сезонное распределение камбалы в Черном море у берегов Грузии. «Природа», 1940, № 4.
Марти Ю. Ю. Промысловая разведка рыбы. Пищепромиздат, 1948.
Марти Ю. Ю. Основные этапы жизненного цикла атлантическо-скандинавских сельдей. Труды ПИНРО. Т. 9, 1956.
Марциновский Е. И. О технике перевозки живых рыб и икры. «Рыбное хозяйство СССР». 1933, № 4.
Медников Б. М. Экологические формы рыб и проблемы симпатрического видообразования. Зоологический журнал. Т. 42. Вып. 1, 1963.
Мейен В. А. Наблюдения над годичными изменениями яичника у окуня. Русский зоологический журнал. Т. VII. Вып. 3, 1927.
Мейен В. А. Очередные задачи методики определения стадий созревания половых продуктов у рыб. Труды Первой Всекаспийской научной рыбохозяйственной конференции. М.-Л., 1936.
Мейен В. А. Инструкция по определению пола и степени зрелости половых продуктов у рыб. Изд. ВНИРО. М., 1939а.
Мейен В. А. К вопросу о годовом цикле изменений яичников костистых рыб. Известия АН СССР, серия биологии, № 3, 19396.
Мейен В. А. О причинах колебаний размеров икринок костистых pыб. Доклады АН СССР. Т. 28, № 7, 1940.
Мейен В. А. Изменение полового цикла самок костистых рыб под влиянием экологических условий. Известия АН СССР. Отдел биологических наук,№ 2, 1944.
Мельянцев В. Г. Форели водоемов Карело-Финской ССР. Петрозоводск, 1952.
Мельянцев В. Г. Палия озер Карело-Финской ССР. Петрозаводск 1953.
Меньшиков М. И. Некоторые данные по рыбам и рыбному хозяйству озер б. Курганского округа Уральской области, 1923.
Меньшиков М. И. К. биологии сибирского осетра (Acipenser baeri) и стерляди (Acipenser ruthenus) p. Иртыша. Ученые записки госуниверситета T. II. Вып. 1, 1936.
Меншуткин В. В. Исследование динамики численности рыб на основе представления популяции в виде кибернетической системы «Вопросы ихтиологии». Т. 4. Вып. 1 (30), 1964.
Месяцев И. И. Запасы рыбы и интенсивность промысле в Северном Каспии. «Рыбное хозяйство», 1935, № 3.
Месяцев И. И. Строение косяков стадных рыб. Известия AНСССР серия биологии, № 3, 1937.
Мешков М. М. Морфологические черты личиночного периода карповых рыб и определительные таблицы их личинок в водоемах Ленинградской области. Ученые записки Ленинградского госуниверситета, серия биологических наук. Вып. 23, 1951.
Мешков М. М. Особенности ранних стадий развития пресноводных рыб. Ученые записки Псковского педагогического института. Вып. 2, 1954.
Милинский Г. И. Биология и промысел морской камбалы (Pleuronectes platessa) Баренцева моря. Труды ПИНРО. Вып. 2, 1938.
Милинский Г. И. Систематика и биология уклей Сямозера. Труды Карело-Финского отделения ВНИОРХ. Т. 2, 1946.
Михайловская А. А. О формообразовании у каспийской морской сельди. Доклады АН СССР. Т. 30, № 6, 1941.
Михеев П. В. Опыт применения искусственных плавучих нерестилищ в дельте р. Дона. «Рыбное хозяйство», 1953, № 7.
Михин В. С. Беломорская и мурманская треска. Известия Отдела прикладной ихтиологии. Т. 3. Вып. 2, 1925.
Михин В. С. Материалы по возрасту и росту аральского сома. Известия Ленинградского научно-исследовательского ихтиологического института. Т. 12. Вып. 1, 1931.
Михин В. С. Материалы по биологии и промыслу угря в восточной части Финского залива. Сборник, посвященный научной деятельности проф. II. М. Книповича. Пищепромиздат, 1939.
Михин В. С. Эффективность естественного нереста семги. Вестник ЛГУ, № 8, 1950.
МихинВ.С. и ШпайхерА.0.0 нерестилищах и естественном нересте семги. Научно-технический бюллетень ВНИОРХ, № 5, 1957.
Мичурин И. В. Сочинения. Т. 1. М. Сельхозгиз, 1948.
Мищенко А. И. Некоторые наблюдения над развитием икры и личинок камбалы. Известия ТИНРО. Т. 14, 1938.
Моисеев П. А. Промысловые камбалы Дальнего Востока. Примиздат, 1946 а.
Моисеев П. А. Некоторые данные о влиянии промысла на состояние стада камбал залива Петра Великого. Зоологический журнал. Т. 25. Вып. 5, 1946б.
Моисеев П. А. Треска и камбалы дальневосточных морей. Труды ПИНРО. Т. 40, 1953.
Мокиевский О. Б. К методике прибрежных гидробиологических исследований. Зоологический журнал. Т. 29. Вып. 4, 1950.
Монастырский Г. Н. Видоизменения доски Э. Леа. Труды научного института рыбного хозяйства. Т. 1, 1924.
Монастырский Г. Н. О методах определения линейного роста рыб До чешуе. Труды научного института рыбного хозяйства. Т. 5, Вып. 4, 1930.
Монастырский Г. Н. Запасы воблы Северного Каспия и методы их оценки. Труды ВНИРО. Т. 11, 1940.
Монастырский Г. Н. Методика оценки состояния запасов и прогно-1U численности северо-каспийской воблы и волжской сельди. Труды ВНИРО. Т, 18, 1951.
Монастырский Г. Н. Динамика численности промысловых рыб. Труды ВНИРО. т. 21, 1952.
Монастырский Г. Н. О типах нерестовых популяций у рыб. Сб. «Очерки по общим вопросам ихтиологии». Изд. АН СССР, 1953.
Мороз В. Н. Размножение рыбца и пути его воспроизводства в условиях зарегулированного стока Днепра. Херсонское книжное и газетное издательство, 1960.
Морозов А. В. Современное состояние запасов воблы Северного Каспия. Бюллетень Всекаспийской экспедиции № 5-6, 1932 а.
Морозов А. В. Несколько соображений о состоянии запасов воблы Северного Каспия. Бюллетень Всекаспийской экспедиции № 1-2, 1932 б.
Морозов А. В. К методике расовых исследований рыб вообще и воблы в частности. Саратов. Нижневолжское краевое изд-во, 1932.
Морозов А. В. К методике установления возрастного состава уловов. Бюллетень океанографического института, № 15, 1934.
Мосевич Н. А. Гидрологические и гидрохимические работы в Волховской губе Ладожского озера в марте 1930 г. Известия Ленинградского ихтиологического института. Т. 12. Вып. 1, 1931.
Москаленко Б. К. Материалы к биологии сиговых рыб Обской губы. Известия ВНИОРХ. Т. 44, 1958.
Мурашкинцева П. А. Биологические группы мурманской сельди по характеру строения зимних колец на чешуе. Труды ПИНРО Т. 1, 1938.
Навозов-Лавров Н. П. Материалы к экологии и промыслу краба в заливе Петра Великого. Производительные силы Дальнего Востока. Т. 4, 1927.
Недошивин А. Я. К биологии осетровых на Дону (из работ Азовской экспедиции). Бюллетень рыбного хозяйства, № 1, 1926.
Недошивин А. Я. Материалы по изучению Донского рыболовства. Труды Азовско-Черноморской научно-промысловой экспедиции. Вып. 4, 1928.
Несмеянов А. Н. Наука – народу. «Природа». 1952, № 4.
Нестеренко Н. В. Опыт гибридизации уральского рипуса с чудским сигом в прудовых условиях. Известия ВНИОРХ. Т. 39, 1957.
Никаноров Ю. И. О популяционной плодовитости европейской ряпушки Coregonus albula (L.) в озерах Латвийской ССР. Доклады АН СССР. Т. 124, № 4, 1959.
Никаноров Ю. И. Отличия в величине, окраске и клейкости икринок у озерной ряпушки-Coregonus albula (L.) различных стад в Латвийской ССР. Зоологический журнал. Т. 38. Вып. 6, 1959.
Никитин В. Н. Планктон Батумской бухты и его годичные изменения. Сборник в честь Н. М. Книповича, 1939.
Никифоров Н. Д. Развитие, рост и выживаемость эмбрионов и молоди семги в естественных условиях. Известия ВННОРХ. Т. 48, 1959.
Николаев И. И. Видовой состав рыб Латвийской ССР Труды Латвийского отд. ВНИРО. Т. 1. 1953.
Николаев И. И. О глубине распространения салаки в Балтийском море. Зоологический журнал. Т. 33. Вып. 3, 1954.
Никольский Г. В. Советская ихтиология за двадцать лет. Зоологический журнал. Т. 17. Вып. 2, 1938.
Никольский Г. В. Рыбы Таджикистана. Изд. АН СССР, 1938.
Никольский Г. В. Инструкция по инвентаризации фауны рыб в заповедниках. Научно-методические записки Главного управления по заповедникам, 1939.
Никольский Г. В. Рыбы Аральского моря. М. Изд. Московского общества испытателей природы, 1940.
Никольский Г. В. Биология рыб. М., 1944.
Никольский Г. В. и др. Рыбы бассейна Верхней Печоры. М., 1947.
Никольский Г. В. Река Амур и ее пресноводные рыбы. Труды Московского общества испытателей природы. Вып. 13. М., 1948.
Никольский Г. В. О биологическом обосновании контингента вылова и путях управления численностью стада рыб. Зоологический журнал. Т. 29. Вып. 1, 1950а.
Никольский Г. В. Амурская ихтиологическая экспедиция 1945- 1949 гг. Труды Амурской ихтиологической экспедиции, 1945-1949 гг. Т. 1. Изд. Моск. общества испытателей природы, 19506.
Никольский Г. В. К систематике пескарей рода Gobio, населяющих бассейн Амура. Бюллетень Московского общества испытателей природы. Т. 55. Вып. 1, 1950в.
Никольский Г. В. О некоторых закономерностях динамики плодовитости рыб. Очерки по общим вопросам ихтиологии. Изд. АН СССР, 1953.
Никольский Г. В. Частная ихтиология. 1954.
Никольский Г. В. Рыбы бассейна Амура. Изд. АН СССР, 1956.
Никольский Г. В. Экология рыб. «Советская наука», 1963.
Никольский Г. В. и Морозова П. Н. О факторах, определяющих величину поголовья стада промысловых рыб Аральского моря. Зоологический журнал. Т. 25. Вып. 4, 1946.
Николюкин Н. И. К методике искусственного рыборазведения. Труды Воронежского отделения Всесоюзного научно-исследовательского института прудового рыбного хозяйства. Т. 1, 1935.
Николюкин Н. И. О гибриде Cyprinus carpio L. X Carassius carassius L. в сравнении с исходными видами. Известия Воронежского педагогического института. Т. III, № 3, 1938.
Николюкин Н. И. Межвидовая гибридизация рыб. 1952.
Николюкин Н. И. и Тимофеева Н. А. Инструкция по выращиванию в прудах осетровых рыб (осетр X стерлядь и белугах стерлядь). М. Изд. Министерства рыбной промышленности, 1956.
Новиков П. И. Результаты вскрытия гнезд беломорской семги на нерестилищах р. Кеми в апреле 1947 г. Бюллетень рыбного хозяйства Карело-Финской ССР, № 1, 1947.
Новиков П. И. Семга р. Кеми. Бюллетень рыбного хозяйства Карело-Финской ССР, № 2, 1948.
Новиков П. И. Об эффективности естественного нереста атлантического лосося. Доклады АН СССР. Т. 18, № 6, 1949.
Новиков П. И. Северный лосось – семга. Биология, промысел и искусственное разведение. Петрозаводск. Изд-во Карело-Финской ССР, 1953.
Овен Л. С. Овогенез и годичный цикл изменений яичников у черноморской султанки Mullus barbatus ponticus Essipov. Труды Карадагской биологической станции АН УССР. Вып. 17, 1961 а.
Овен Л. С. О специфике икрометания и о плодовитости черноморской султанки Mullus barbatus ponticus Essipov. «Вопросы ихтиологии». Вып. 17, 1961 б.
Очерки по общим вопросам ихтиологии. Изд. АН СССР, 1953.
Очерки по биологическим основам рыбного хозяйства (Вопросы теории динамики численности рыб). Изд. АН СССР, 1961.
Павловский Е. Н. О необходимости развития систематики. Зоологический журнал. Т. 31. Вып. 2, 1952.
Павловский Е. Н. Содержание ихтиологии и положение ее среди сопредельных наук. Очерки по общим вопросам ихтиологии. Изд. АН СССР,
1953.
Парин Н. В. Летучие рыбы северо-западной части Тихого океана. Труды Института океанологии. Т. 31, 1960.
Певзнер В. И. Песчанки Баренцева моря. Труды научно-исследовательского института рыбного хозяйства. Т. III. Вып. 2, 1928.
Пентегов Б. П., Ментов Ю. Н. и Курнаев Е. Ф. Физико-химическая характеристика нерестово-миграционного голодания кеты. Известия Тихоокеанской научно-промысловой станции. Т. 2. Вып. 1, 1928.
Перцева Т. А. Нерест каспийских сельдей в Северном Каспии. Труды ВНИРО. Т. 14, 1940.
Пераева-Остроумова Т. А. Систематика икринок и предличинок сельдевых северной части Каспийского моря. Труды ВНИРО. Т. 18, 1951.
Петров В. В. К систематике русских корюшек. Известия Отдела прикладной ихтиологии. Т. 3. Вып. 1, 1925.
Петров В. В. К познанию кавказских уклеек. Известия Бакинской ихтиологической лаборатории. Т. 2, Вып. 1, 1926.
Петров В. В. Материалы по систематике русских корюшек. Ладожская корюшка. Известия Отдела прикладной ихтиологии. Т. 4. Вып. 1, 1926.
Петров В. В. Материалы по изучению роста и возраста каспийских осетровых. Известия Отдела прикладной ихтиологии. Т. 6. Вып. 2, 1927.
Петров В. В. Материалы по росту и возрасту азовского сазана. Труды Азовско-Черноморской научно-промысловой экспедиции. Вып. 8, 1931.
Петров В. В. К характеристике сазана оз. Иссык-Куль. Труды Киргизской комплексной экспедиции. АН СССР. Т. III. Вып. 2, 1935.
Петров В. В. и Петрушевский Г. К. Материалы по структуре чешуи у сазана (Cyprinus carpio). Известия Отдела прикладной ихтиологии. Т. 9. Вып. 3.1929.
Пирожников П. Л. Инструкция по сбору и обработке материалов по питанию рыб. Изд. ГосНИОРХ, 1953.
Поведение рыб и промысловая разведка (Сборник статей) Труды ВНИРО. Т. 36, М., 1958.
Подлесный А. В. Белорыбица Stenodus leucichtys Quid. Биоэкологический очерк. Труды Сибирского отделения ВНИОРХ. Т. 7. Вып. 1, 1947.
Пожалуева Е. В. Материалы по возрасту и темпу роста аральского сазана. Труды научного института рыбного хозяйства. Т. 3. Вып. 2, 1928.
Покровский В. В. К систематике окуня. Труды Ленинградского общества естествоиспытателей. Т. 60. Вып. 1, 1930.
Покровский В. В. Материалы по исследованию внутривидовой изменчивости окуня (Perca fluviatilis L.). Труды Карело-финского отделения ВНИОРХ. Т. 3, 1951.
Покровский В. В. Ряпушка озер Карело-Финской ССР. Петрозаводск, 1953.
Покровский В. В. и Смирнов А. Ф. Очерк рыболовства Онежского озера. «Рыбное хозяйство Карелии». Вып. 1, 1932.
Покровский В. В. и Беляева К. И. О балхашском губаче. Ученые записки Карело-Финского университета. Т. II (1947). Вып. 3, 1948.
Покровский В. В. и Новиков П. И. Озера Карелии и их рыбохозяйственное значение. «Озера Карелии». Гос. изд-во Карельской ССР, Петрозаводск, 1959.
Померанцев Г. П. К методике обратного расчисления длин и весов рыбы. Труды Уральского отделения ВНИОРХ. Т. 4, 1949.
Попов А. М. Кефали (Mugilidae) Европы. Труды Севастопольской биологической станции АН СССР. Т. 2, 1930.
Попова В. П. Характеристика состояния запасов камбалы Черного моря в 1955 г. и возможный вылов на 1956 г. Аннотация к работам, выполненным ВНИРО в 1955 г., Сб. 1, 1956.
Попова В. П. Характеристика состояния запаса камбалы (Rhombus maeoticus) в 1956 г. Аннотация к работам, выполненным ВНИРО в 1956г., Сб.II, 1958.
Попова К. С. Влияние голодания на развитие кутума в начале личиночного периода жизни. Труды Института морфологии животных им. Северцева. Вып.33,1961.
Потапова О. И. Лещ как объект озерного рыбного хозяйства Карело-Финской ССР. Карело-Финский филиал АН СССР. Материалы совещания по проблеме повышения рыбной продуктивности водоемов Карело-Финской ССР. 1954.
Потапова О. И. Условия размножения леща в Миккельском озере. Материалы по повышению рыбной продуктивности малых Озер Карелии. Труды Карельского филиала АН СССР. Т. II, 1956.
Правдин И. Ф. Наблюдения над каспийской миногой (Caspiomyzon wagneri Kessler) весной 1912 года. Труды Астраханской ихтиологической лаборатории. Т. 2, Вып. 6, 1913.
Правдин И. Ф. Описание некоторых форм русской плотвы. Материалы русского рыболовства. Т. 4. Вып. 9, 1915.
Правдин И. Ф. Биометрическая характеристика промысловых признаков волховского сига. Материалы по исследованию Волхова и его бассейна. Вып. III. Ч. I, 1924а.
Правдин И. Ф. Описание некоторых форм русской плотвы. Плотва пестовская. Известия Отдела прикладной ихтиологии. Т. 2, 19246.
Правдин И. Ф. Схема измерений рыб семейства осетровых. Известия Государственного института опытной агрономии. Т. 2. Вып. 3. 1924 в.
Правдин И. Ф. Один из приемов, облегчающий работу по систематике рыб. Русский гидробиологический журнал № 7-9, 1925.
Правдин И. Ф. Очерк западнокамчатского рыболовства. Известия Тихоокеанской научно-промысловой станции. Т. 1. Вып. 1, 1928.
Правдин И. Ф. Амурская горбуша Oncorhynchusgorbuscha natio amurensis. Известия ВНИОРХ. Т. 14, 1932.
Правдин И. Ф. Указания к составлению научно-промысловых карт. Известия ВНИОРХ. Т. 18-19, 1934.
Правдин И. Ф. Руководство по изучению рыб. Изд. Ленинградского государственного университета, Л., 1939.
Правдин И. Ф. Система рыбообразных и рыб. (О кн. Л. С. Берга- Система рыбообразных и рыб, ныне живущих и ископаемых). Научный бюллетень Ленинградского государственного университета, № 2, 1945.
Правдин И. Ф. Проблема однолетнего выращивания сазана (Cyprinus carpio L.) в пойменных водоемах. Ученые записки Ленинградского университета № 75. Серия биологии. Вып. 15, 1945.
Правдин И. Ф. Научно-промысловые карты. Труды Первой научно-технической конференции по рыбной промышленности. Карело-Финской ССР. Петрозаводск, 1947.
Правдин И. Ф. К материалам по миграции ладожского лосося. Известия Карело-Финской базы АН СССР, № 2, 1948.
Правдин И. Ф. Русская ихтиология за последние 30 лет (1917- 1947). Ученые записки Карело-Финского университета. Т. 3. Вып. 3, биологические науки,1949.
Правдин И. Ф. Вопросы методики ихтиологических исследований. Схемы определения счетных и пластических признаков рыб. Известия Карело-Финского филиала АН СССР. Вып. 1, 1951.
Правдин И. Ф. Сиги водоемов Карело-Финской ССР. М.-Л., Изд. АН СССР, 1954.
Правдин И. Ф. Проблема повышения рыбной продуктивности внутренних водоемов Карело-Финской ССР. Материалы Совещания по проблеме повышения рыбной продуктивности внутренних водоемов Карело-Финской ССР, 1954.
Правдин И. Ф. Вопросы методики ихтиологических исследований. 4. Методы определения плодовитости рыб. Труды Карельского филиала АН СССР. Вып. 13, 1958.
Правдин И. Ф. Вопросы методики ихтиологических исследований. Методы исследования миграций рыб. Труды Карельского филиала АН СССР. Вып. 13, 1958.
Привольнев Т. И. Методы изучения развития рыб. Жизнь пресных вод СССР. Т. 4. Ч. 2, 1959.
Пробатов А. Н. О возрасте аральского сома. Известия Отдела прикладной ихтиологии. Т. 9. Вып. 2, 1929.
Пробатов А. Н. Материалы по изучению осетровых рыб Амура. Ученые записки Пермского госуниверситета. Т. 1, Вып. 1, 193u.
Программы и наставления для наблюдения и собирания коллекций по естественной истории. 7-ое изд., 1913.
Рабинерсон А. И. О систематическом положении беломорской сельди. Известия Госинститута опытной агрономии. Вып. 2, 1923.
Рабинерсон А. И. Материалы по исследованию беломорской сельди. Труды научно-исследовательского института по изучению Севера. Вып. 25, 1925.
Рабинерсон А. И. Исследования над беломорской и мурманской сельдью летом 1925 г. Известия Отдела прикладной ихтиологии. Т. 4. Вып. 1, 1926.
Рабинерсон А. И. Замечания по поводу статьи С. Аверинцева «Сельди Белого моря». Ч. 1. Известия Отдела прикладной ихтиологии. Т, 6. Вып. 1, 1927.
Рабинерсон А. И. Определение роста рыб и связанные с ним вопросы. «Природа», 1927, № 4.
Рабинерсон А. И. Характеристика уловов сельди залива Петра Великого весною 1926 г. Известия Отдела прикладной ихтиологии. Т. 8, 1928.
Расс Т. С. Работы группы по изучению икры и мальков (ихтиопланктона). Доклад первой сессии Государственного океанографического института (14-22 апреля 1931). М., 1931.
Расс Т. С. Инструкция по сбору и обработке сборов икринок и мальков рыб. 1935.
Расс Т. С. Исследование ранних стадий развития некоторых костистых рыб Баренцева моря. ВНИРО. 1939 а.
Расс Т. С Инструкция по сбору икринок и мальков рыб. Пищепромиздат, 19396.
Расс Т. С О размножении и жизненном цикле мурманской сельди (Clupea harengus harengus L.). Труды ПИНРО, Вып. 6, 1939 в.
Расс Т. С Ступени онтогенеза костистых рыб (Teleostei). «Зоологический журнал», Т. 25, Вып. 11, 1946.
Расс Т. С. Значение исследований размножения рыб для оценки возможных уловов. «Рыбное хозяйство», 1953, № 2.
Расс Т. С и Казанова И. И. Инструкция по сбору икринок и личинок рыб. Изд. ВНИРО. М., 1958.
Рахманова С. И. О мойве из Баренцева моря. Труды научного института рыбного хозяйства. Т. III. Вып. 2, 1928.
Редкозубов Ю. Н. Видоизменения доски Монастырского для обратного расчисления темпа роста рыб по чешуе. Труды Карело-Финского отделения ВНИОРХ. Т. 3, 1951.
Редкозубов Ю. Н. Микропроектор из микроскопа МБИ (МБР-1). «Вопросы ихтиологии». Т. 4. Вып. 3, 1964.
Романов Н. С. Указатель литературы по рыбному хозяйству южных бассейнов СССР за 1918-1953 гг. (сырьевая база и воспроизводство запасов промысловых рыб). М., Изд. АН СССР, 1955.
Романов Н. С. Аннотированный указатель опубликованных в 1956 г. работ по проблеме «Закономерности динамики численности, поведения и распределения рыб, морских млекопитающих, промысловых беспозвоночных и водорослей в связи с условиями существования». «Рыбное хозяйство». М., 1959.
Романов Н. С. Указатель литературы по рыбному хозяйству Дальнего Востока за 1923-1956 гг. Изд-во АН СССР, 1959.
Романовский В. И. О статистических критериях принадлежности данной особи к одному из близких видов. Труды Туркестанского научного общества при Среднеазиатском госуниверситете. Т. 2, 1925.
Ромейс Б. Микроскопическая техника. Изд-во «Иностранная литература». М., 1953.
Роскин Г. И. Микроскопическая техника. Изд-во «Советская наука». М., 1951.
Руденко Г. П. Возрастной состав рыб, ихтиомасса и рыбопродуктивность окуневых озер. Научно-технический бюллетень ГосНИОРХ, № 16, 1962.
руководство по изучению питания рыб в естественных условиях. Изд. АН СССР, 1961.
Руководство по методике исследований физиологии рыб. Изд. АН СССР, 1962.
0’Рурк. Таблицы умножения. М.-Л., ОНТИ, 1937.
Рыбы Белого моря. Карельское книжное изд-во, 1958.
Рыженко М. П. Личинки и мальки мурманской сельди. Сборник в честь Н. М. Книповича, 1939.
Сабанеев Л. П. Описание и изображение рыб, встречающихся в Российской империи. СПб, 1875.
Савельев В. И. Выращивание стандартного сеголетка в северных районах СССР. Известия ВНИОРХ. Т. 24, 1941.
Савина Н.О. и Покровский В.В. Посезонное распределение промысловых рыб в озере Балхаше. Известия ВНИОРХ. Т. 37, 1956.
Сальников Н. И. и Малятский С. М. К систематике белуги Азово-Черноморского бассейна. Труды научной рыбохозяйственной и биологической станции Грузин. Т. 1. Вып. 1, 1934.
Сафьянова Т. Е. Мечение ставриды в Черном море. Труды Азчер-НИРО. Вып. 17, 1958.
Сборник статей по биохимии рыб (перевод с англ.) Изд. иностр. литературы, 1953.
Сборник статей по методике определения возраста и роста рыб. Красноярск, 1926.
Световидов А. Н. Материалы по систематике и биологии хариусов озера Байкала. Труды Байкальской лимнологической станции. Т. 1, 1931.
Световидов А. Н. Европейско-азиатские хариусы (genus Thymallus Cuvier). Труды Зоологического института АН СССР. Т. III, 1936.
Световидов А. Н. Трескообразные. Фауна СССР. Рыбы. Т. 9. Вып. 4, 1948.
Световидов А. Н. Сельдевые (Clupeidae). Фауна СССР. Рыбы. Т. 2. Вып. I, 1952.
Световидов А. Н. Рыбы Черного моря. «Наука», 1964.
Себенцов Б. М. и Михеев П. В. Рыбоводный эффект искусственных плавучих нерестилищ в технических водохранилищах. «Рыбное хозяйство», 1949. № 2.
Северцов С. А. Динамика населения и приспособительная эволюция животных. Изд. АН СССР, 1941.
Смирнов А. Г. Возраст и рост аральской шемаи. Известия Отдела прикладной ихтиологии. Т. 9. Вып. 2, 1929.
Смирнов А. Н. Запасы и прогнозы улова азовской хамсы на 1937- 1938 гг. «Рыбное хозяйство», 1937, № 10-11.
Смирнова К. В. Нельма оз. Зайсан. Известия АН СССР, серия зоологии; № 5, 1945.
Соин С. Г. Об особенностях биологии размножения европейского и амурского сомов. Зоологический журнал. Т. 57, № 6, 1947.
Соин С. Г. Эмбрионально-личиночный период развития верхогляда. «Вопросы ихтиологии». Вып. 13, 1959.
Соколовская И. И. Преципитиновая реакция в гибридизации. Известия АН СССР. Отдел математики и естественных наук, серия биологии, № 2- 3,1936.
Соколовская И. И. Серологические исследования при отдаленной гибридизации. Известия АН СССР. Отдел математики и естественных наук, № 4, 1938.
Солдатов В. К. Исследование биологии лососевых Амура. Т. 1. СПб, 1912.
Солдатов В. К. Исследование осетровых Амура. Материалы к познанию русского рыболовства. Т. III. Вып. 12, 1915.
Солдатов В. К. Рыбы реки Печоры. Труды Северной научно-промысловой экспедиции. Вып. 17, 1924.
Солдатов В. К. и Линдберг Г. У. Обзор рыб дальневосточных морей. Известия ТИНРО. Т. 17, 1930.
Соловьева Н. С. Время закладки зимних колец и начала прироста по чешуе сельди в различных районах Баренцева моря. Труды ПИНРО. Вып. 2, 1938.
Сонина М. А. Треска Белого моря. Сб. «Материалы по комплексному изучению Белого моря». Т. 1, 1957.
Стеллер. Сочинение Стеллера «Из Камчатки в Америку». Неопубликованные работы хранятся в Архиве АН СССР, 1928.
Строганов Н. С. Экологическая физиология рыб. Изд. МГУ, 1962.
Суворов Е. К. Этюды по изучению каспийских сельдей. Труды Каспийской экспедиции 1904 г. Т. 1, 1907.
Суворов Е. К. К ихтиофауне Балтийского моря. Тр. Балтийской экспедиции. Вып. 2, 1913.
Суворов Е. К. К ихтиофауне Чешской губы. Труды Института по изучению Севера. Вып. 43, 1929.
Суворов Е. К. и др. Биология мойвы. Труды ихтиологического научно-исследовательского института. Т. И, 1931.
Суворов Е. К. Использование скрытых возможностей роста рыб. Информационный сборник консультационного бюро ВНИОРХ, № 4, 1940.
Суворов Е. К. К вопросу о расширении рыбосырьевой базы Финского залива. Записки ЛГУ, 1945.
Суворов Е. К. Основы ихтиологии. Изд-во «Советская наука», 1948.
Суворов Е. К. Промысловые водоемы СССР, Изд. ЛГУ, 1948.
Суворов Е. К., Вадова Л. А., Сынкова А. И. Биометрические измерения мойвы. Труды Ленинградского научно-исследовательского института рыбного хозяйства. Т. 3. Вып. 1, 1931.
Суворкина М. И. Темп роста двух форм ладожских сигов Coregonus lavaretus baeri n. ladagae и С. lavaretus baeri n. swirensis. Труды Петергофского биологического института. Т. 12, 1934.
Сушкин П. П. и Белинг Д. Е. Определитель рыб пресноводных и морских Европейской России. Пг, 1923.
Сыроватский И. Я. К биологии размножения донской сельди. «Природа», 1939, №5.
Сыч-Аверинцева Н. В. О Pleuronectes flesus Баренцева и Белого морей. Труды Института рыбного хозяйства. Т. V. Вып. 4, 1930.
Талиев Д. Н. К познанию трески Белого моря. Известия Ленинградского научно-исследовательского ихтиологического института. Т. II. Вып. 2, 1931.
Талиев Д. Н. и Базикалова А. Я. Предварительные результаты сравнений фауны Байкала и Каспия при помощи реакции преципитации. Доклады АН СССР, Т. 2, № 8, 1934.
Талиев Д. Н. К познанию реакции гемоагглютинации у рыб. Труды Байкальской лимнологической станции, № 1, 1935.
Талиев Д. Н. К изучению остеологии байкальских Cottoidei при помощи лучей Рентгена. Вестник рентгенологии. Т. 20, 1938.
Талиев Д. Н. Опыт применения реакции преципитации к познанию происхождения и истории Байкальской фауны. Труды Байкальской лимнологической станции. Т. 10, 1940.
Талиев Д. Н. Серологический анализ рас байкальского омуля. Труды Зоологического института АН СССР. Т. 6. Вып. 4, 1941.
Талиев Д. Н. Новый род Cottoidei из оз. Байкала. Доклады АН СССР. Т.54,№ 1, 1946 а.
Талиев Д. Н. Предки байкальских Cottoidei в Ципо-Ципиканских озерах. Доклады АН СССР. Т. 52. № 8, 1946 б.
Талиев Д. Н. Серологический анализ некоторых диких и одомашненных форм сазана. (Cyprinus carpio L.). Труды Зоологического института АН СССР. Т. 8. Вып. 1, 1946в.
Талиев Д. Н. Бычки-подкаменщики Байкала (Cottoidei). М.-Л., Изд-во АН СССР, 1955.
Танасийчук В. С. Молодь воблы. Труды ВНИРО. Т. II. 1940.
Танасийчук В. С. Учет промысловых рыб в северном Каспии. Труды ВНИРО. Т. 18, 1951.
Танасийчук Н. П. Нерестовые миграции волжской многотычинковой сельди. Труды Волго-Каспийской рыбохозяйственной станции. Т. 10. Вып. 1, 1948.
Танасийчук Н. П. Промысловые рыбы Волго-Каспия. М. Пищепромиздат, 1951.
Танфильев Г. И. Моря: Каспийское, Черное, Балтийское, Ледовитое, Сибирское и Восточный океан. 1931.
Таранец А. Я. Краткий определитель рыб советского Дальнего Востока и прилегающих вод. Известия ТИНРО. Т. 11, 1937.
Телегин К. Ф. Зимнее рыболовство Сорокского района, в 1928- 1949 гг. Известия Ленинградского ихтиологического института Т. 11. Вып. 2, 1932.
Телегин К. Ф. Как увеличить уловы салаки. «Рыбное хозяйство» 1938, № 12.
Телегин К. Ф. Промысловая разведка салаки на Финском заливе. Информационный бюллетень консультационного бюро ВНИОРХ, № 3, 1939.
Телегин К. Ф. О флюгерном лове салаки. Информац. бюллетень консультационного бюро ВНИОРХ, № 2, 1939.
Терещенко 3. П. Материалы по биологии и промыслу каспийской кефали. Труды Каспийского бассейнового филиала ВНИРО. Т. XI, 1950.
Терещенко К. К. Лещ Каспийско – Волжского района, его промысел и биология. Труды Астраханской ихтиологической лаборатории. Т .4. Вып. 2, 1917.
Титенков И. С. Жилая нельма в Кубенском озере. «Рыбное хозяйство», 1951, № 10.
Титенков И. С. Кубенская нельма. М., Изд-во «Рыбное хозяйство», 1961.
Тихий М. И. Методика механического вычисления роста рыб. Труды Петроградского общества естествоиспытателей. Т. 48. Вып. 1, 1917.
Тихий М. И. Западно-Камчатская горбуша и ее возраст. Известия Отдела прикладной ихтиологии. Т. 4. Вып. 2, 1926.
Тихий М. И. Гидроэлектрические станции и рыболовство на реке Свири. Известия Отдела прикладной ихтиологии. Т. VI. Вып. 1, 1927.
Тихий М. И Наблюдения над икрометанием весенненерестующих рыб. Известия ВНИОРХ. Т. 21, 1939.
Тихонов В. Н О систематическом положении мурманской сельди. Труды ПИНРО. Вып. 7, 1941.
Тихонов В. Н. Зимнее распределение сельди (Clupea harengus harengus) в южной части Баренцева моря. Труды ПИНРО. Т. 4, 1939.
Травин В. И. Новый вид морского окуня в Баренцевом море (Sebastes mentella Travin sp. nov.). Доклады АН СССР. Т. 77. № 4, 1951.
Тремпович П. В. Дифференцированная окраска чешуи промысловых рыб для определения возраста. Бюллетень Всекаспийской научно-рыбохозяйственной экспедиции № 3-4, 1932.
Третьяков Д. К. Сейсмосензорные каналы сельдевых. Зоологический журнал. Т. 17. Вып. 5, 1938.
Третьяков Д. К. Очерки по филогении рыб. 1944.
Троицкий С. К. О рыбах в бассейне реки Кубани. «Природа», 1939, № 10.
Троицкий С. К. Рыбы Краснодарского края. Краснодар, 1948.
Трошин А. С. Метод радиоактивных индикаторов и его применение в гидробиологии. В кн. «Жизнь пресных вод СССР». Т. 4. Ч. 1, 1956.
Труды Всесоюзной конференции по вопросам рыбного хозяйства. Изд. АН СССР, 1953.
Труды совещания по проблеме акклиматизации рыб и кормовых беспозвоночных. Изд. АН СССР, 1954.
Труды Совещания по методике изучения кормовой базы и питания рыб. Изд. АН СССР, 1955.
Труды совещания по динамике численности рыб. Изд. АН СССР, 1961.
Трусов В. 3. Гистологический анализ так называемой IV стадии зрелости яичников судака. Труды лаборатории основ рыбоводства. Т. 1, 1947.
Трусов В. 3. Годичный цикл яичников донского судака и особенности отдельных моментов цикла у судака других водоемов. Труды лаборатории основ рыбоводства. Т. II, 1949 а.
Трусов В. 3. Озимая и яровая расы осетровых. Доклады АН СССР. Т. 67, № 3, 1949 б.
Турдаков Ф.А. Рыбы Киргизии, Фрунзе, 1952.
Тюрин П. В. К вопросу изучения азиатской корюшки. Труды Сибирской ихтиологической лаборатории. Т. 2. Вып. 1, 1924.
Тюрин П. В. О зависимости между длиной рыбы и ее весом. Труды Сибирской ихтиологической лаборатории. Т. 2. Вып. 3. 1927.
Тюрин П. В. Биологические основания реконструкции рыбных запасов в северо-западных озерах СССР. Известия ВНИОРХ. Т. 11, 1957.
Тюрин П. В. Определение возможностей рыбопродуктивности озер северо-западной части СССР. «Вопросы ихтиологии». Вып. 13, 1959.
Тюрин П. В. Фактор естественной смертности рыб и его значение при регулировании рыболовства. «Вопросы ихтиологии». Т. 2. Вып. 3 (24), 1962.
Тюрин П. В. Биологические обоснования регулирования рыболовства на внутренних водоемах. Пищепромиздат, 1963,
Федоров П. Ф. Материалы по биологии и промыслу беломорской корюшки. Известия Ленинградского ихтиологического института. Т. II. Вып. 2, 1931.
Феклистова М. В. Об эколого-географической изменчивости рыб. Известия Карело-Финского филиала АН СССР, № 1, 1951.
Филатов Д. П. и Дуплаков С. Н. Материалы к изучению рыб Аральского моря. Бюллетень Среднеазиатского государственного университета, № 14, 1926.
Филатова 3. А. Количественный учет донной фауны юго-западной части Баренцева моря. Труды ПИНРО. Т. II, 1938.
Филипченко Ю. А. Изменчивость и методы ее изучения. Л., 1929.
Фортунатов М.А. Форели Севанского озера. Concpecies Saimo isch-chan Kessler. Ч. 1. Систематика. Труды Севанской озерной станции. Т. 1. Вып. 2, 1927.
Фортунатова К. Р. Методика изучения питания хищных рыб. I. Зоологический журнал. Т. 30. Вып. 6, 1951.
Фортунатова К. Р. Методика изучения питания хищных рыб. II. Труды Совещания по методике изучения кормовой базы и питания рыб. Изд. АН СССР, 1955.
Фрейдлинг В. А. Гидрологическая характеристика озер Микнельского, Крошнозера. Труды Карельского филиала АН СССР, II, 1956.
Фролов А. И. О локальных формах сахалинской сельди. Труды ТИНРО. Т. 32, 1950.
Xаилов К. М. Электрофоретическое исследование белков крови тресковых рыб. Труды Мурманского морского биологического института. Вып. 4/8, 1962.
Xалдинова Н. А. Материалы по размножению и развитию беломорской наваги. Зоологический журнал. Т. 15. Вып. 2, 1936.
Чернов В. К. Материалы по биологии хариуса. Труды Бородинской биологической станции. Вып. 2, 1934.
Чернавин В. Я. Опыт систематической группировки некоторых Salmonoidei, основанный на остеологических признаках. Известия Гос. института опытной агрономии. Т. 2, 1923.
Черновский А. А. Население рек, озер и их продуктивность. Озера Карелии. Изд. Бородинской биологической станции. Петрозаводск, 1930.
Черфас Б. И. Рыбоводство в естественных водоемах. М. Пищепромиздат, 1956.
Чугунов Н. Л. Определение возраста и темпа роста рыб по костям. Сборник статей по методике определения возраста и роста рыб. Красноярск, 1926.
Чугунов Н. Л. Биология молоди промысловых рыб Волго-Каспийского района. Труды Астраханской научно -рыбохозяйственной станции. Т. 6. Вып. 4, 1928.
Чугунов Н.Л. Опыт биостатического определения запасов рыб в Северном Каспии. «Рыбное хозяйство», 1935, № 6 и 8.
Чугунова Н. И. Методика изучения возраста большеглазого пузанка. Труды ВНИРО. Т. 14, 1940 а.
Чугунова Н. И. Рост осетровых Азовского моря. «Рыбное хозяйство», 19406, № 5.
Чугунова Н. И. Распределение бычков в Северном Каспии. Зоологический журнал. Т. 25. Вып. 5, 1946.
Чугунова Н. И. Методика изучения возраста и роста рыб. Изд-во «Советская наука». М., 1952.
Чугунова Н. И. Руководство по изучению возраста и роста рыб. Изд. АН СССР, 1959.
Шапошникова Г. X. Биология молоди семги и тинды реки Выг. Труды Карельской научно-исследовательской станции. Т. 1, 1935.
Шапошникова Г X. Изучение ихтиофауны водоемов. Изд. АН СССР, 1950.
Шерешевская Е. Г. Заболевание рыб и рыбьей икры сапролегнией. «Рыбное хозяйство Карелии». Вып. 1, 1932.
Шеханова И. А. Методика массового мечения молоди осетровых рыб радиоактивным фосфором. Труды Совещания по физиологии рыб. 1958.
Шмальгаузен И. И. Определение основных понятий и методика исследования роста. Рост животных. Сборник работ. М.-Л., 1935.
Шмидт П. Ю. Рыбы восточных морей Российской империи. СПб, 1904.
Шмидт П. Ю. Миграции рыб. М.-Л., Изд-во АН СССР, 1947.
Шмидт П. Ю. Рыбы Тихого океана. М., Пищепромиздат, 1948.
Шмидт П. Ю. Рыбы Охотского моря. М.-Л., Изд-во АН СССР, 1950.
Шорыгин А. А. Промысел кильки в Каспийском море и перспективы его развития. Рыбная промышленность СССР. Сборник 1-2, 1947.
Шорыгин А. А. Количественный способ изучения пищевой конкуренции у рыб. «Зоологический журнал». Т. 25. Вып. 1, 1946.
Шорыгин А. А. Годовая динамика пищевой конкуренции у рыб. Зоологический журнал. Т. 27. Вып. 1, 1948.
Шорыгин А. А. Питание и пищевые взаимоотношения рыб Каспийского моря. М., Пищепромиздат, 1952.
Шуколюков А. М. Рост молоди рыб из низовьев р. Урала. Известия ВНИОРХ. Т. 14, 1932.
Щербинин И. А. К методике обработки содержимого кишечника некоторых рыб. «Вопросы ихтиологии». Вып. 4, 1955.
Энгельс Ф. Диалектика природы. 1948.
Юданов И. Г. Условия нереста и развития икры ряпушки в заморной зоне Обской губы. «Рыбное хозяйство», 1939, ,№ 4.
Юровицкий Ю. Г. Некоторые Вопросы Методики изучения плодовитости рыб. «Вопросы ихтиологии». Вып. 10, 1958.
Antipa Gr, Die Clupeinen des westlichen Teiles des Schwarzen Meeres und der Donoumundungen. Denkschr. Akad. Wiss. Wien, math-naturw. Kl., LXXYIII, 1905.
Вауег Н. A method of measuring fish eggs. Proceed. 4 Intern. Fish. Congress Washington 1908. Bull. U. S. Bureau of Fisherei’s, XXYIII, part 2 (1910).
Berg L. S. Obersicht der Verbreitung Susserwasserfische Europas. Zoo-geografick, V. I., 1932.
Gushing I. and Sprague I. Agglutination of eritrocytes of various Fishes by human and others Sera Amer. Naturalist, v. 87, 1953.
Сushing I. and Duva11 G. Isoagglutination in the fishes. Amer. Naturalist, v. 21, 1957.
Davidson F. A. a Shostrom О. Е. Physikal and chemical changes the pine Salmon during the spawning migration. Report, № 33 Bureau of Fisheries, 1936.
Drihion A. Etude biologique de quelques protides seriques de Sangs de prissons du moyen de 1’electrophorese sur papier C. R. Soc. Biol., 148, 13-14:
1218-1220, 1954. ‘
Duncker G. Variation und Vermandschaft von Pleuronectes flesus und Pt. platessa untersucht mittelst der Heinsk’schen Methode, 1896.
Ehrenbaum. Der Flussnal, 1930.
Eiсhwa1d E. Faunae Caspii maris primitae. Bull. Soc. Nat. Moscou, XI, 1838.
Farran G. Fluctuation in the stock of Herring an the North Coast of Donegal. Rap. Pr. Verb. LXY, 1930.
Fowler. The marine fishes of west Africa, 1936.
Fulton T. W. On the rate of growth of Fishes. The Twenty fourth Annual Report of the Fishery Board of Scotland, 1906, VIII.
Gemeroy D. G. On the relationship of some common fishes as determined by the precipition resction. Zoological Society, v. 28, part, 3, N, 15-19, pp. 109-124, 1943.
Georguiev J. et Kolarov P. Sur la repartition Hivernale et les migrations du saurel Ie long du Littoral Bulgare, 1956.
Gilbert Ch. Age at maturity of the Pacific coast Salmon of the genus Oncorhynchus. Bull. U. S. Bur. Fish., XXXII, 1912.
Gоs1ine W. The sensory canals the head in some Cyprinodont fishes with particular reference to the genus Fundulus. Occasional papers of the Museum Zoology, № 519, 1949.
Heckel J. und Kner R. Die Susswasserfische der Oestreichischen Mo-narchie. Leipzig, 1858.
Heinсke. Naturgeschichte des Herings, 1898.
Hicks R. and Litt1e C. The Blood Relationship of four of mice. genetics, v. 16, № 5, p. 397, 1931.
HodgsonW. The Forecasting of the rast England Herring Fishery. Journ. of Anim. Ecol., v. V, n. 2, 1932.
Irwin M. and Cole L. Immunogenetic Studies of Species and of Species Hubrids in Doves and the Separation of Species Substances in the Backoross. Journ. Exp. Zool., vol. 73, № 1, 1936 a.
Irwin M. and Cole L. Immunogenetic Studies of Species Hubrids from the Cross Columba livia and Streptopelia risoria. Journ. Exp; Zool., v. 73, № 2, 1936 b.
Jaaskelainen.Om fiskarna och fisket i Ladoga. Finlands Fiske-rier, IV, 1917.
Jarvi. Uber die Arten und Formen der Corgonen in Finland, 1928.
Jordan D. A classification of Fishes. Stanf. Univ. Publ. Biol. Sci, .III (2), 1923.
Jordan D. and Ekerman B.W.A review of the giant Mackerellike Fishes Tunnies, Spearfishes and Swordfishes Oc. pap. Calif. Acad. Sci. XII, 1926.
Journal duconseil, v. VII, № 1, 1932, v. XIX, № 2, 1953.
Joshida Joichi. A way of marking fish specimens with their original body-colours kept. Bull. Misaki Marine Biol. Inst. Kyoto Univ., № 3, 1962.
Keyvanfar A. Serologic et immunologie de deux especes de thonides (germo alaliinga gmelin et Thunnus thunnus iinue) de 1’Atlantique et de la mediterranec. Revue des travaux de 1’inestituts des peches maritimes Decembre, 1962.
Коdama H. Die Differenzierung des Kaviars vor anderen Fischrogen Arch. f. Hug., Bd. 78, 1913.
Koelz W. Coregonid fishes of the Great Lakes. Bull. Bur. Fish., XLIII, p. II, 1927 (1929).
Kulmatycki. Studien an Coregonen Polens. Arch. liydrobiol. i rybactwa, I (1926), Suwaiki II (1927).
К у chick i L. Swinka (Chondrostoma nasus), 1931.
Landsteiner A. Serological tests with the blood of cavia porcelilus and cavia rufercens proc. Soc. exp. biol. a. Med., vol. 28, p. 981, 1931.
Lea E. On the Methods used in the Hering investigation, 1910.
Linnaeus C. Systema naturae. Ed. X., 1758.
Litynski. Sieja i sielawa w jeziorach suwalskich i augustowskich. Sprawozd. Stac. Hydrobiol. na Wigrach, № 2-3, 1923.
Marukawa H. Biological und Fishery of Japanese Kingcrab Paialitho-des camtshatica, 1933.
Meek A. The migrations of Fish. London, 1916.
Neresheimer E. Ueber den Nachweis der Blutverwandtschaft bei Fischen durch die Serumdiagnose. Allg. Fisch-Ztg., jg. 33, № 24, 1908.
Neresheimer E. Blutverwandtschaft und Serumrcaktionen bei Salmo-niden, Ber. a. d. Bayr. Biol. Vers-Stat. in Munden, Bd. 2, Stuttgart, 1909.
Normann I. A systematic Monograph of the Flatfishes, 1934.
Otterstrom. Helfting (Coregonus albula L) og Helf (Coregonus lavaretus L.) i Danmakr. Kolding, 1922.
Pallas P. S. Zoographia ross-asiatica, 111. Petropoli, 1811.
Raunsefell G. A. and Everhart H. W. Fishery Science its Methods and Applications. New-York and London, 1953.
O’Rourke F.J. Serological relations in the genus gabus Natura, v. 183, №4669, pp. 1192,1959.
O’Rourke F. J. Seroligical criteria of Species Fish. C. Med. Sei., v. 410, 1960.
Reibisсh. Uber den Einfluss der Temperatur auf die Entwickelung von Fischeiern, 1902.
RidgwayG., Gushing I. Serological differentiation of population of Sockey Salmon (onerca) fish. and Wildlife ser. Sci., Rep. fish, № 257, 1958.
Ridgway G., dishing I. and D u v a 11. Serological differentiation of population of Sockey Salmon (onerca) fish. and Wildlife ser. Sci., Rep. fish., № 257, 1958.
Ridgway G., Clontz G. and Matsumoto C. Intraspecifie differences in the Serum antigens of Red Salmon, demonstrated by immu-nochemical methods. Bureau of Commercial Fisheries Seattle, Washington, 1959.
Ridgway G. and Clontz G. Blood types in pacific Salmon. Spec. Sci. Papers. Fisheries, 324, 1960.
Rо11efsen G. The Spawning Zone in Cod Otoliths and Prognosis of Stock. Rep. Norw. Fish. a. Marine Inv. IV, № 11, 1935.
Schaperclaus W. Fischereiwirtscheftslehre (seen und Flusswirtschaft)-B книге Fischerei Kufde. Radabe und Berlin, 1953.
Sheriff G. Report on the mathematical analysis of random samples of herrings. Fisherey Boani Юг Scotland, Sc. Investigations, N. I, Edinburgh. 6, 1922.
Sindermann С. and Meirs D. A major blood group system in at-lantic Sea herring. Copia, № 3, 1959.
Schmidt Joh. Contributions to the life history of the eel (Anguilla vulgaris FL). Rapp. Proc- Verb. Cons. Perm Int. Expl. Mer. v. 5, 1906.
Schmidt Joh Die Laichplatze des Flussaales. Inter. Rev. d. ges. Hyd-robiol. u. Hydrogr., Bd. XI, 1923.
Smitt F. Kritisk forteckn’ng ofver de i Riksmuseum befintliga Salmoni-der. K. Svenska Vet. Akad. Handlingar, № 8, 1886.
Steinmann P. Monogfaphie der schweizerichen Coregonen. Revue Suisse d’Hydroligie, XII, f. 2, 19p0.
Suzuki A., Morio T. d Mimоtо К– Serological studies of the races of tuna. II rept. Manttai, Red. Fish. Lab. v. II, 1959. .
Swingle H. and Smith E. Management of Ponds for the Production of Game and Pan FisP- A Symposium of Hydrobiology. Madison, 1941.
Thienemann A. Die Unterschiede zwieschen der grossen Marane-des Madusees und des Selenters-ees. Zool. Anz., v. 48, 1916.
Thienemann A. Biologische Forschungsreisen und das System der Bio-logie. Leipzig, г., 1920.
Thienemann A. Weitere Untersuchungen an Coregonen. Arch. f. Hydrobiol., XIII, 1922.
Thienemann A. Coregonen aus dem Ladogasee. Vanamon Julkaisuja, v. I, № 7, 1927.
Thienemann A. Uber die Eddmarasane (Coregonus lavaretus forma generosis Pet.), 1928.
Tomijama A. Gobiidae of Japan. Japanese journal of Zoology № 1, 1936.
Val1e К. Suomen kalat, 1934.
|
ПРИЛОЖЕНИЕ 1 |
||||||||||
|
Таблица промеров рыб семейства карповых(Cyprinidae) |
||||||||||
|
Признаки |
Номер рыбы |
|||||||||
|
Пол и стадия зрелости Вес рыбы, г Возраст Длина всей рыбы (аb или L), мм* Длина по Смитту (ос) Длина рыбы без С (ad или l) Длина туловища (od) Формула боковой линии Чешуи по боку хвостового стебля Лучей в D Лучей в А Формула глоточных зубов Тычинок на первой жаберной дуге Позвонков (туловищных и хвостовых) Ось тела Длина рыла (an) Диаметр глаза (горизонтальный) (пр) Заглазничный отдел головы (ро) Длина головы (ао) Высота головы у затылка (lт) Ширина лба Наибольшая высота тела (gh) Наименьшая высота тела (ik) Антедорсальное расстояние (aq) Постдорсальное расстояние (rd.) Длина хвостового стебля (fd) Длина основания D (qs) Наибольшая высота D (tu) Длина основания А (yy1) Наибольшая высота А (ej) Длина Р (vx) Длина V (zz1) Расстояние между Р и V (vz) Расстояние между V и А (гу) |
||||||||||
|
* Все измерения в этом и последующих приложениях даются в мм, |
||||||||||
ПРИЛОЖЕНИЕ II
Ведомость индексов признаков рыб семейства карповых
(Cyprinidae)
|
Признаки |
Номер рыбы |
|||||||||
|
Пол и стадия зрелости Вес рыбы, г Возраст Длина рыбы без С (ad или l) Чешуи в боковой линии Лучей в D Лучей в Л Тычинок на первой жаберной дуге Позвонков (туловищных и хвостовых) В % длины телаДлина рыла Диаметр глаза Заглазничный отдел головы Длина головы Высота головы у затылка Ширина лба Наибольшая вые эта тела Наименьшая высота тела Антедорсальное расстояние Постдорсальное расстояние Длина хвостового стебля Длина основания D Наибольшая высота D Длина основания Л Наибольшая высота Л Длина Р Длина V Расстояние между Р и V Расстояние между V и Л В % длины головы Длина рыла Диаметр глаза Ширина лба |
||||||||||
ПРИЛОЖЕНИЕ III
Таблица промеров рыб семейства лососевых (Salmonidae)
|
Признаки |
Номер рыбы |
|||||||||
|
Пол и стадия зрелости Вес рыбы, г Возраст Длина всей рыбы (аb или L), мм* Длина по Смитту (ос) Длина рыбы без С (ad или l) Длина туловища (od) Формула боковой линии Чешуи по боку хвостового стебля Лучей в D Лучей в А Формула глоточных зубов Тычинок на первой жаберной дуге Позвонков (туловищных и хвостовых) Ось тела Длина рыла (an) Диаметр глаза (горизонтальный) (пр) Заглазничный отдел головы (ро) Длина головы (ао) Высота головы у затылка (lт) Ширина лба Наибольшая высота тела (gh) Наименьшая высота тела (ik) Антедорсальное расстояние (aq) Постдорсальное расстояние (rd) Длина хвостового стебля (fd) Длина основания D (qs) Наибольшая высота D (tu) Длина основания А (yy1) Наибольшая высота А (ej) Длина Р (vx) Длина V (zz1) Расстояние между Р и V (vz) Расстояние между V и А (гу) |
||||||||||
ПРИЛОЖЕНИЕ IV
Ведомость индексов признаков рыб семейства лососевых (Salmonidae)
|
Признаки |
Номер рыбы |
|||||||||
|
Пол и стадия зрелости Вес рыбы, г Возраст Длина по Смитту (ас) Чешуи в боковой линии Лучей в D Лучей в А Тычинок на первой жаберной дуге Пилорических придатков Позвонков (туловищных и хвостовых) В % длины тела (по Смитту) Длина рыла Диаметр глаза Заглазничный отдел головы Длина средней части головы Длина головы Высота головы у затылка Ширина лба Длина верхнечелюстной кости Длина нижней челюсти Наибольшая высота тела Наименьшая высота тела Антедорсальное расстояние Постдорсальное расстояние Антевентральное расстояние Антеанальное расстояние Длина хвэстовм’0 стебля Длина основания D Наибольшая высота D Длина основания А Наибольшая высота А Длина Р Длина V Расстояние между Р и V Расстояние между V и А В % длины головы Длина рыла Диаметр глаза Заглазничный отдел головы Длина средней части головы Высота готовы у затылка Ширина лба Длина верхнечелюстной кости Ширина верхнечелюстной кости Длина нижней челюсти Высота рыльной площадки Ширина рыльной площадки Наименьшая высота тела |
||||||||||
|
Признаки |
Номер рыбы |
|||||||||
|
Высота рыльной площадки в % ее длины Диаметр глаза в % длины рыла Ширина верхнечелюстной кости в % ее длины Наименьшая высота тела в % длины нижней челюсти Расстояние V–A в % антедорсального расстояния Расстояние V-A в % расстояния P–V Длина V в % длины Р Длина жаберной тычинки в % длины жаберной дуги |
||||||||||
ПРИЛОЖЕНИЕ V
Таблица промеров рыб семейства осетровых (Acipenseridae)
|
Признаки |
Номер рыбы |
|||||||||
|
Пол и стадия зрелости Вес рыбы, г Длина всей рыбы (ab или L) Длина рыбы до конца средних лучей С (ас) Длина до корней средних лучей С (ad или l) Длина туловища (od.) Спинных жучек Боковых жучек Брюшных жучек Лучей в D Лучей в А Тычинок на первой жаберной дуге Длина рыла (an) Ширина рыла Расстояние от конца рыла до губы Расстояние от конца рыла до хрящевого свода рта (1-2) Расстояние от конца рыла до средних усиков (1-3) Расстояние от основания усиков до хрящевого свода рта Длина наибольшего усика (4-5) Ширина рта (6-7) Диаметр глаза (горизонтальный) (пр) 3аглазничный отдел головы (ро) Длина головы (ао) Высота головы у затылка (lт) Ширина головы на вертикали середины головы Наибольшая ширина головы |
||||||||||
|
Признаки |
Номер рыбы |
|||||||||
|
Ширина лба Высота лба Наибольшая высота тела (qh) Наибольшая толщина тела Наибольший обхват тела Наименьшая высота тела (ik) Длина хвостового стебля (fd) Антедорсальное расстояние (aq) Антевентральное расстояние (аг) Антеанальное расстояние (ау) Длина основания D Наибольшая высота D (tu) Длина основания A (yy1) Наибольшая высота A (ej) Длина Р (vх) Длина V (zz1) Расстояние между Р и V (vz) Расстояние между V и Л (гу) |
||||||||||
ПРИЛОЖЕНИЕ VI
Таблица промеров рыб семейства сельдевых (Clupeidae)
|
Признаки |
Номер рыбы |
|||||||||||||||
|
Пол и стадия зрелости Вес рыбы, г Возраст Длина всей рыбы (аb или L) Длина рыбы до конца средних лучей С (ас) Длина рыбы без С (ad или l) Поперечных рядов чешуи Лучей в D Лучей в А Тычинок на первой жаберной дуге Пилорических придатков Позвонков (отдельно туловищных и хвостовых) Брюшных шипиксв впереди ануса Брюшных шипиков позади ануса Зубов на верхней челюсти Зубов на нижней челюсти Зубов на сошнике Ось тела Длина рыла (an) Диаметр глаза (вертикальный) (n1p1) Заглазничный отдел головы (ро) Длина головы (ао) Высота головы у затылка (lт) Ширина лба |
||||||||||||||||
|
Продолжение |
||||||||||||||||
|
Признаки |
Номер рыбы |
|||||||||||||||
|
Длина верхнечелюстной кости (а1а6) Длина нижней челюсти (a1l1) Наибольшая высота тела (qh) Наименьшая высота тела (ik) Антедорсальное расстояние (aq) Постдорсальное расстояние (rd) Антевентральное расстояние (az) Антеанальное расстояние (ау) Длина хвостового стебля (fd) Длина основания D (qs) Наибольшая высота D (tu) Длина осноования A (yy1) Наибольшая высота A (ej) Длина Р (vх) Длина V (zz1) Расстояние между Р и V (vг) Расстояние между V и A(zу) Длина средних лучей С (dc) |
||||||||||||||||
ПРИЛОЖЕНИЕ VII
Таблица промеров окуневых (Percidae) и скорпеновых (Scorpaenidae)
|
Признаки |
Номер рыбы |
|||||||||
|
Пол и стадия зрелости Вес рыбы, г Возраст Длина всей рыбы (ab) Длина рыбы без С (ad) Чешуи в боковой линии (l.l.) Позвонков (туловищных и хвостовых) Лучей в ID Лучей в IID Лучей в А Тычинок на первой жаберной дуге Длина первой жаберной дуга Длина наибольшей жаберной тычинки Длина головы (ао) Высота головы (lm) Длина рыла (an) Диаметр глаза (горизонтальный) (np) Заглазничный отдел головы (ро) Длина верхнечелюстной кости (a1,a2) Длина нижнечелюстной кости (k1,l1) Ширина лба Наибольшая высота тела (qh) |
||||||||||
|
Продолжение |
||||||||||
|
Признаки |
Номер рыбы |
|||||||||
|
Наименьшая высота тела (ik) Длина хвостового, стебля (fd) Антедорсальное расстояние (aq) Расстояние от ануса до A (Ау) Длина основания ID (qs) Длина основания IID (q1s1) Расстояние между ID и IID Высота ID (tu) Высота IID (t1u1) Длина Р (vx) Ширина Р (vx1) Длина V (zz1) Длина основания А (уу1) Высота A (ej) Хвостовая выемка (cb) |
||||||||||
ПРИЛОЖЕНИЕ VIII
Ведомость индексов признаков рыб семейства окуневых (Percidae) и скорпеновых (Scorpaenidae)
|
Признаки |
Номер рыбы |
||||||||||
|
Пол и стадия зрелости Вес рыбы, г Длина тела Длина головы В % длины тела Длина головы Высота головы у затылка Наибольшая высота тела Наименьшая высота тела Длина хвостового стебля Антедорсальное расстояние Постдорсальное расстояние Расстояние от ануса до А Длина основания ID Длина основания IID Промежуток между ID и IID Высота ID Высота IID Длина основания А Высота А В % длины головы Длина рыла Диаметр глаза горизонтальный Заглазничный отдел головы Длина верхнечелюстной кости Длина нижнечелюстной кости |
|||||||||||
|
Продолжение |
|||||||||||
|
Признаки |
Номер рыбы |
||||||||||
|
Ширина лба Длина головы в % наибольшей высоты тела Длина жаберной тычинки в % длины жаберной дуги Ширина основания Р в % длины Р Хвостовая выемка |
|||||||||||
ПРИЛОЖЕНИЕ IX.
Таблица промеров миноговых (Petromyzonidae)
|
Признаки |
Номер рыбы |
|||||||||||||||||||||||||||
|
Пол и стадия зрелости Вес, г Вся длина (аbили L) Наибольшая высота тела (qh) Наибольший обхват тела Высота головы (lт) Расстояние от конца рыла до первого жаберного отверстия (ае) Длина жаберного аппарата Длина рыла (an) Диаметр глаза (пр) Промежуток между глазом и жабгрным аппаратом (рe) Диаметр ротового диска (ik) Длина головы с жаберным аппаратом (ао) Ширина лба Антедорсальное расстояние (aq) Расстояние от конца рыла до ануса (аА) Промежуток между ID и IID (sq1) Длина основания ID (qs) Наибольшая высота ID (t u) Длина основания IID (q1s1) Наибольшая высота IID (t 1u1) Длина спинной части С (s1b) Расстояние от ануса до конца С (Аb) Зубов на верхнечелюстной пластинке Зубов на нижнечелюстной пластинке Зубов на язычной пластинке |
||||||||||||||||||||||||||||
|
ПРИЛОЖЕНИЕ X |
||||||||||||||||||||||||||||
|
Таблица квадратов чисел от 0,01 до 1,99 |
||||||||||||||||||||||||||||
|
N |
N2 |
N |
N2 |
N |
N2 |
N |
N2 |
N |
N2 |
N |
N2 |
|||||||||||||||||
|
0,01 0,02 0,03 0,04 0,05 0,06 0,07 0,08 0,09 0,10 0,11 0,12 0,13 0,14 0,15 0,16 0,17 0,18 0,19 0,20 0,21 0,22 0,23 0,24 0,25 0 26 0,27 0,28 0,29 0,30 0,31 0,32 0,33 0,34 |
0,0001 004 009 016 025 036 049 064 081 100 121 144 1?9 196 225 256 289 324 361 400 441 484 529 576 625 676 729 784 841 900 961 0,1024 089 156 |
0,35 0,36 0,37 0,38 0,29 0,40 0,41 0,42 0,43 0,44 0,45 0,46 0,47 0,48 0,49 0,50 0,51 0,52 0,53 0,54 0,55 0,56 0,57 0,58 0,E9 0,60 0,61 0,62 0,63 0,64 0,65 0,66 0,67 0,68 |
225 296 369 444 521 600 681 764 849 936 0,2025 116 2C9 304 401 500 601 704 8C9 916 0,3025 136 249 364 481 600 721 844 969 0,4096 225 356 489 624 |
0,69 0,70 0,71 0,72 0,73 0,74 0,75 0,76 0,77 0,78 0,79 0,80 0,81 0,82 0,83 0,84 0,85 0,86 0,87 0,88 0,89 0,90 0,91 0,92 0,93 0,94 0,95 0,96 0,97 0,98 0,99 1,00 1,01 1,02 |
761 0,4900 0,504 184 329 476 625 776 929 0,6084 241 400 561 724 889 0,7056 225 296 519 744 921 0,8100 281 464 649 83b 0,9025 216 409 ?04 801 1,0000 201 404 |
1,03 1,04 1,05 1,06 1,07 1,08 1,09 1,10 1,11 1,12 1,13 1,14 1,15 1,16 1,17 1,18 1,19 1,20 1,21 1,22 1,23 1,24 1,25 1,26 1,27 1,28 1,29 1,30 1,31 1,32 1,33 1,34 1,35 1,36 |
609 816 1,1025 236 449 664 881 1,2100 321 544 769 996 1,3225 456 689 924 1,4161 400 641 884 1,5129 376 625 876 1,6129 384 ?41 900 1,7161 424 689 956 1,8225 496 |
1,37 1,38 1,39 1,40 1,41 1,42 1,43 1,44 1,45 1,46 1,47 1,48 1,49 1,50 1,51 1 52 1,53 1,54 1,55 1,56 1,57 1,58 1,59 1,60 1,61 1 62 1,63 1,64 1,65 1,66 1,67 1,68 1,?9 1,70 |
7?9 1,9944 321 600 881 2,0164 449 736 2,1025 316 609 904 2,2201 500 801 2,3104 409 716 2,4025 336 649 9c4 2,5281 600 921 2,6244 569 896 2,7225 556 889 2,8224 561 2,8900 |
1,71 1,72 1,73 1,74 1,75 1,76 1,77 1,78 1,79 1,80 1,81 1,82 1,83 1,84 1,85 1,86 1,87 1,88 1,89 1,90 1,91 1,92 1 93 1,94 1,95 1,93 1,97 1,98 1,99 |
2,9241 584 929 3,0276 625 976 3,1329 684 3, 2041 400 761 3,3124 489 856 3,4225 596 969 3,5344 721 3,6100 481 864 3,7249 639 3,8025 416 809 3,9204 601 |
|||||||||||||||||
|
ПРИЛОЖЕНИЕ XI |
||||||||||||||||||||||||||||
|
Квадратные корни из чисел |
||||||||||||||||||||||||||||
|
0 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
|||||||||||||||||||
|
0,0 0,1 0,2 0,3 0,4 0,5 0,6 |
0,0000 3162 4472 5477 6325 7071 7746 |
1000 3317 4583 5568 6403 7141 7810 |
1414 3464 4690 5657 6481 7211 7874 |
1732 3606 4795 5745 6557 7280 7937 |
2000 3742 4899 5831 6633 7248 8000 |
2236 3873 5000 5916 6708 7416 8062 |
2449 4000 5099 6000 6782 7483 8124 |
2646 4123 5193 6083 6856 7550 8185 |
2828 4243 5292 6164 6928 7616 8246 |
3000 4359 5385 6245 7000 7681 8307 |
||||||||||||||||||
|
Продолжение прилож. XI |
||||||||||||||||||||||
|
0 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
|||||||||||||
|
0,7 0,8 0,9 1,0 1,1 1,2 1,3 1,4 1,5 1,6 1,7 1,8 1,9 2,0 2,1 2,2 2,3 2,4 2,5 2,6 2,7 2,8 2,9 3,0 3,1 3,2 3,3 3,4 3,5 3,6 3,7 3,8 3,9 4,0 4,1 4,2 4,3 4,4 4,5 4,6 4,7 4,8 4,9 5,0 5,1 5,2 5,3 5,4 5,5 5,6 5,7 5,8 |
8367 8944 9487 1 0000 0488 0954 1402 1832 2247 2649 3038 3416 3784 4142 4491 4832 5166 5492 5811 6125 6432 6733 7029 7321 7607 7889 8166 8439 8708 8974 9235 9494 9748 2,0000 0248 0494 0736 0976 1213 1448 1679 1909 2136 2361 2,2583 2804 3022 3238 3452 3664 3875 4083 |
8426 9000 9539 0050 0536 1000 1446 1874 2288 2689 3077 3454 3820 4177 4526 4866 5199 5524 5893 6155 6462 6763 7059 7349 7635 7916 8193 8466 8735 9000 9261 9519 9774 0025 0273 0518 0761 1000 1237 1471 1703 1932 2159 2383 2605 2825 3043 3259 3473 3685 3896 4104 |
8485 9Э55 9592 0100 0583 1045 1489 1916 2329 2728 3115 3491 3856 4213 4560 4900 5232 5556 5875 6186 6492 6793 7088 7378 7664 7914 8221 8493 8762 9026 9287 9545 9799 0050 0298 0543 0785 1024 1260 1494 1726 1954 2181 2405 2627 2847 3065 3281 3495 3707 3917 4125 |
8544 9110 9344 0149 0630 1091 1533 1958 2369 2767 3153 3528 3892 4248 4595 4933 5264 5588 5906 6227 6523 6823 7117 7407 7692 7972 8248 8520 8788 9053 9313 9570 9824 0075 0322 0567 0809 1048 1284 1517 1749 1977 2204 2428 2650 2869 3087 3302 3516 3728 3937 4145 |
8602 9165 9695 0198 0677 1136 1576 2000 2410 2806 3191 3565 3928 4283 4629 4956 5297 5620 5937 6248 6553 6852 7146 7436 7720 8000 8276 8547 8815 9079 9339 9596 9849 0100 0347 0591 0833 1071 1307 1541 1772 2000 2226 2450 2672 2891 3108 3324 3537 3749 3958 4166 |
8660 9220 9747 0247 0724 1180 1619 2042 2450 2845 3229 3601 3964 4318 4663 5000 5320 5652 5959 6279 6583 6882 7176 7464 7748 8028 8303 8574 8841 9105 9365 9621 9875 0125 0372 0616 0857 1095 1331 1564 1794 2023 2249 2472 2694 2913 3130 3345 3558 3770 3979 4187 |
8718 9274 9798 0296 0770 1225 1662 2083 2490 2884 3266 3638 4000 4353 4697 5033 5362 5684 6000 6310 6613 6912 7205 7493 7776 8055 8330 8601 8868 9131 9391 9 547 9900 0149 0396 0640 0881 1119 1354 1587 1817 2045 2271 2494 2716 2935 3152 3367 3580 3791 4000 4207 |
8775 9327 9849 0344 0817 12?9 1705 2124 2530 2923 3304 3675 4036 4387 4731 5067 5395 5716 6031 6340 6643 6941 7234 7521 7804 8083 8358 8628 8894 9157 9416 9672 9925 0174 0421 0664 0905 1142 1378 1610 1840 2068 2S93 2517 2738 2956 3173 3388 3601 3812 4021 4228 |
8832 9381 9899 0392 0863 1314 1747 2166 2570 2961 3342 3711 4071 4422 4765 5100 5427 5748 6062 6371 6673 6971 7263 7550 7833 8111 8385 8655 8921 9183 9442 9698 9950 0199 0445 0688 0928 1666 1401 1633 1863 2091 2316 2539 2760 2978 3195 34G9 3622 3833 4042 4249 |
8888 9434 9950 0440 0909 1358 1790 2207 2610 3000 3379 3748 4107 4457 4799 5133 5460 5780 6093 6401 6703 7000 7292 7578 7861 8138 8412 8682 8947 9209 94.58 9723 9975 0224 0469 0712 0952 1190 1424 1656 1886 2113 2338 2561 2782 3000 3216 3431 3643 3854 4062 4269 |
||||||||||||
|
5.9 6.0 6.1 6.2 6.3 6.4 6.5 6.6 6.7 6.8 6.9 7.0 7.1 7.2 7.3 7.4 7.5 7.6 7.7 7.8 7.9 8.0 8.1 8.2 8.3 8.4 8.5 8.6 8.7 8.8 8.9 9.0 9.1 9.2 9.3 9.4 9.5 9.6 9.7 9.8 9.9 10.0 11,0 12,0 13,0 14,0 15,0 16,0 17,0 18,0 19,0 20,0 |
4290 4495 4698 4900 5100 5S93 5495 5690 5884 6077 6268 6458 6646 6833 7019 7203 7386 7568 7749 7928 8107 8284 8460 8636 8810 8983 9155 9326 9496 9665 9833 3,0000 0166 0332 0496 06?9 0822 0984 1145 1305 1464 1623 3166 4641 6056 7417 8730 4,0000 1231 2426 3589 4721 |
4310 4515 4718 4920 5120 5318 5515 5710 5904 6096 6287 6476 6665 6851 7037 7221 7404 7586 7767 7946 8125 8302 8478 8653 8827 9000 9172 9343 9513 9682 9850 0017 0183 0348 0512 0676 0838 1000 1161 1321 1480 1780 3317 4785 6194 7550 8859 0125 1352 2644 3704 4833 |
4331 4536 4739 4940 5140 5338 5534 5729 5923 6115 6306 6495 6683 6870 7055 7240 7423 7604 7785 7964 8142 8320 8496 8671 8844 9017 9189 9360 9530 9698 9866 0033 0199 0364 0529 0692 0854 1016 1177 1337 1496 1937 3466 4928 6332 7683 8987 0249 1473 2661 3818 4944 |
4352 4556 4759 4960 5159 5357 5554 5749 5942 6134 6325 6514 6702 6889 7074 7258 7441 7622 7703 7982 8160 8337 8513 8688 8862 9034 9206 9377 9547 9715 9883 0050 0216 0381 0545 0708 0871 1032 1193 1353 1512 2С94 3615 5071 6469 7815 9115 0373 1593 2778 3932 5056 |
4372 4576 4779 4980 5179 5377 5573 5768 5962 6153 6344 6533 6721 6907 7092 7276 7459 7641 7821 8000 8178 8355 8531 8705 8879 9052 9223 9394 9563 9732 9900 0067 0232 0397 0561 0725 0887 1048 1209 1369 1528 2249 3764 5214 6606 7947 9243 0497 1713 2895 4045 5166 |
4393 4597 4799 5000 5199 5S97 5Е93 5788 5981 6173 6363 6552 6739 6926 7111 7295 7477 7659 7839 8018 8195 8373 8548 8723 8893 9019 9240 9411 9580 9744 9917 0083 0249 0414 0578 0741 0903 1064 1225 1385 1544 2404 3912 5355 6742 8079 9370 0620 1833 3012 4159 5277 |
4413 4617 4819 5020 5219 ?417 56125807 6000 6192 6382 6571 6758 6944 7129 7313 7495 7677 7857 8036 8213 8390 8566 8740 8914 9086 9257 9428 9597 9766 9933 0100 0265 0430 0594 0757 0919 1081 1241 1401 1559 2558 4059 5496 6878 8210 9497 0743 1952 3128 4272 5387 |
4434 4637 4839 5040 5239 ?436 5632 5826 6019 6211 6401 6589 6777 6963 7148 7331 7514 7695 7875 8054 8231 8408 8583 8758 8931 9103 9275 9445 9614 9783 9950 0116 0282 0447 0610 0773 0935 1097 1257 1417 1575 2711 4205 5637 7014 8341 9623 0866 2071 3243 4385 5497 |
4454 4658 4860 5060 5259 5456 5652 5845 6038 6230 6420 6608 6796 6981 7166 7350 7532 7713 7893 8071 8249 8425 8601 8775 8948 9120 9292 9462 9631 9799 9967 0133 0299 0463 0627 0790 C952 1113 1273 1432 1?91 2863 4351 5777 7148 8471 9749 0988 2190 3359 4497 5607 |
4474 4678 4880 5080 5278 5475 5671 5865 6058 6249 6439 6627 6814 7000 7185 7368 7550 7731 7911 8089 8267 8443 8618 8792 8965 9138 9309 9479 9648 9816 9983 0150 0315 0480 0643 0806 0968 1129 1289 1448 1607 3015 4496 5917 7283 8601 9875 1110 2308 3474 4609 5715 |
||||||||||||
Оглавление
Предисловие……………………………………………………………………………………………………………… 5
Введение……………………………………………………………………………………………………………………. 7
Глава I. Основная литература по методике ихтиологических исследований и рыбам водоемов СССР…………………………………………………………………………………………………………………………….. 11
Глава II. Исследования систематики рыб…………………………………………………………………. 17
Таксономические единицы и правила научной номенклатуры ……………………………… 17
Диагностическое значение меристических и пластических признаков……………………. 22
Схемы описания рыб разных семейств……………………………………………………………………. 27
Значение в систематике рыб анатомических признаков …………………………………………. 82
Применение иммуносерологических реакций в ихтиологии …………………………………… 87
Глава III. Пол и половая зрелость…………………………………………………………………………….. 95
Характеристика соотношения полов……………………………………………………………………….. 93
Стадии зрелости гонад и сравнительная оценка отдельных шкал зрелости ………….. 98
Коэффициенты и индексы зрелости……………………………………………………………………….. 107
Сбор и фиксирование гонад …………………………………………………………………………………… 109
Глава IV. Определение плодовитости рыб …………………………………………………………….. 110
Значение изучения плодовитости рыб…………………………………………………………………… 110
Индивидуальная плодовитость …………………………………………………………………………….. 111
Относительная плодовитость………………………………………………………………………………… 115
Рабочая плодовитость …………………………………………………………………………………………… 115
Плодовитость порционно нерестующих рыб…………………………………………………………. 116
Видовая плодовитость …………………………………………………………………………………………… 118
Популяционная плодовитость……………………………………………………………………………….. 119
Глава V. Нерест и нерестилища………………………………………………………………………………. 122
Общие указания о наблюдениях над нерестом рыб……………………………………………….. 122
Установление сроков и продолжительности нереста ……………………………………………. 125
Определение температурного порога нереста………………………………………………………… 126
Процессы откладывания и оплодотворения икры………………………………………………… 127
Характер нерестилищ ……………………………………………………………………………………………. 128
Плотность кладок икры у лососей…………………………………………………………………………. 133
Исследования озерных нерестилищ ………………………………………………………………………. 134
Орудия для сбора икры………………………………………………………………………………………….. 137
Способы учета отложенной и оплодотворенной икры…………………………………………… 139
Искусственные нерестилища ………………………………………………………………………………… 140
Глава VI. Изучение жизненных циклов и молоди рыб ………………………………………….. 142
Изучение жизненных циклов рыб …………………………………………………………………………. 142
Изучение молоди рыб……………………………………………………………………………………………… 145
Орудия лова молоди рыб……………………………………………………………………………………….. 155
Глава VII. Определение возраста рыб…………………………………………………………………….. 163
Значение исследований возраста рыб ……………………………………………………………………. 163
Определение возраста по чешуе……………………………………………………………………………… 168
Определение возраста по костям и отолитам…………………………………………………………. 179
Глина VIII. Определение темпа роста рыб…………………………………………………………….. 192
Метод и приборы для обратного расчисления темпа роста…………………………………… 192
Определение возрастного состава рыб в уловах …………………………………………………… 211
Средние размеры некоторых рыб по возрастным группам ………………………………….. 212
Методы определения скрытых (потенциальных) возможностей роста рыб ………… 214
Определение упитанности и жирности рыб……………………………………………………………. 219
Глава IX. Миграции ……………………………………………………………………………………………….. 222
Классификация миграций и их биологическое значение……………………………………….. 222
Методы изучения миграций…………………………………………………………………………………… 227
Способы мечения рыб и образцы меток……………………………………………………………….. 231
Обработка результатов мечения……………………………………………………………………………. 245
Глава X. Учет состояния рыбных запасов и определение возможных уловов………. 247
Краткий обзор методов определения рыбных запасов и прогнозирования уловов. 247
Примеры определения фактической численности рыб, ихтиомассы и возможных уловов в обработанных химическим методом озерах……………………………………………………………….. 265
Математическое моделирование динамики численности рыб………………………………. 268
Глава XI. Принципы составления рыбопромысловых карт………………………………….. 271
Определение термина и содержания научно-промысловой карты……………………….. 271
Картирование общегеографических сведений о водоеме……………………………………… 273
Картирование гидрофизических элементов………………………………………………………….. 277
Картирование гидрохимических элементов…………………………………………………………… 282
Картирование гидробиологических элементов……………………………………………………… 284
Картирование ихтиологических данных ………………………………………………………………. 287
Картирование сведений о современном промысле ……………………………………………….. 295
Сведения о перспективах хозяйства………………………………………………………………………. 297
Глава XII. Применение вариационной статистики в ихтиологических работах……. 300
Значение индексов и модальной варианты……………………………………………………………. 301
Вариационные ряды………………………………………………………………………………………………. 301
Средняя величина (М) и ее нахождение…………………………………………………………………. 304
Среднее квадратическое или основное уклонение ?. Нахождение ? способом уклонения и способом сумм…………………………………………………………………………………………………………….. 313
Коэффициент вариации С и средние ошибки М, ? и С ………………………………………….. 314
Дифференция рядов……………………………………………………………………………………………….. 321
Коэффициент корреляции………………………………………………………………………………………. 327
Коэффициент регрессии R………………………………………………………………………………………. 332
Общие замечания к вариационно-статистическим работам………………………………….. 334
Литература …………………………………………………………………………………………………………….. 336
ПРИЛОЖЕНИЯ
Приложение I. Таблица промеров рыб семейства карповых (Cyprinidae)……………. 363
Приложение II. Ведомость индексов признаков рыб семейства карповых
(Cyprinidae)……………………………………………………………………………………………………………… 364
Приложение III. Таблица промеров рыб семейства лососевых (Salmonidae)………… 465
Приложение IV. Ведомость индексов признаков рыб семейства лососевых
(Salmonidae)…………………………………………………………………………………………………………….. 366
Приложение V. Таблица промеров рыб семейства осетровых (Acipenscridae)……… 367
Приложение VI. Таблица промеров рыб семейства сельдевых (Clupeidae) 368 Приложение VII. Таблица промеров окуневых (Percidae) и скорпеновых
(Scorpaenidae)…………………………………………………………………………………………………………. 369
Приложение VIII. Ведомость индексов признаков рыб семейства окуневых (Percidae) и скорпеновых (Scorpaenidae) ……………………………………………………………………………………….. 370
Приложение IX. Таблица промеров миноговых (Petromyzonidae)…………………………. 371
Приложение X. Таблица квадратов чисел от 0,01 до 1,99………………………………………. 372
Приложение XI. Квадратные корни из чисел………………………………………………………… 372
Leave a Reply
Оставьте первый комментарий!